theperthgroup.com/SCIPAPERS/EPENatBioTech1993.pdf
theperthgroup.com/SCIPAPERS/biotek8.html
virusmyth.com/aids/hiv/epwbtest.htm
Is a Positive Western Blot Proof of HIV Infection?
Eleni Papadopulos-Eleopulos, Valendar F. Turner y John M. Papadimitriou
Bio/technology 11, 696-707 (1993).
"...we are not simply contending in order that my view or that of yours may prevail, but I presume we ought both of us to be fighting for the truth...".
from Philebus, the Dialogues of Plato.
Resumen
Se acepta en la actualidad que un test de anticuerpos de VIH Western blot (WB) positivo es sinónimo de infección por VIH, con el consiguiente riesgo de desarrollar y morir de sida. En este artículo, presentamos una evaluación crítica de los datos disponibles en la actualidad respecto al aislamiento del VIH y los tests de anticuerpos. Los datos disponibles indican que: (I) los tests de anticuerpos no están estandarizados; (II) los tests de anticuerpos no son reproducibles; (III) las proteínas del WB (bandas) que se consideran que están codificadas por el genoma del VIH y que son específicas al VIH podrían no estar codificadas por el genoma del VIH, y podrían en verdad representar proteínas celulares normales; (IV) incluso si las proteínas fuesen específicas al VIH, puesto que no se ha utilizado ningún estándar oro para determinar la especificidad, y podría incluso no existir, un WB positivo podría representar únicamente una reactividad cruzada con los muchos anticuerpos que no son del VIH presentes en los pacientes de sida y aquellos en riesgo de sida, y de este modo no estar relacionado con la presencia del VIH. Concluimos que el uso de los tests de anticuerpos del VIH como una herramienta diagnóstica y epidemiológica de la infección de VIH debería reconsiderarse.
Introducción
En la actualidad, los únicos métodos utilizados rutinariamente para demostrar la presencia del VIH in vivo son los tests ELISA y WB. En el ELISA, las "proteínas del VIH" se presentan como una mezcla, mientras que en el WB las proteínas del VIH se disocian y se colocan en una tira de gel de poliacrilamida. Tras la electroforesis, que separa las proteínas por su peso molecular y su carga, las proteínas se transfieren a una membrana de celulosa mediante electroblotting. En el desarrollo del test de anticuerpos, tanto del ELISA como del WB, el suero del paciente se añade a la preparación de antígenos. Se supone que si los anticuerpos del VIH están presentes, reaccionarán con las proteínas del VIH, y tras un lavado se visualiza mediante una reacción cromógena de inmunoglobulina antihumana de enzimas. En el ELISA, la reacción se lee ópticamente. Para el WB, las proteínas individuales se reconocen e interpretan visualmente como bandas coloreadas, cada una de las cuales se designa con una pequeña "p" (de proteína), seguida por un número (que es el peso molecular en kilodaltons), por ejemplo la p41.
Se cree que el WB es altamente sensitivo y específico, considerándose un resultado positivo como sinónimo de infección por VIH. Un estado VIH positivo tiene unas implicaciones tan profundas y de largo alcance que nadie debería llevar esta carga sin sólidas garantías acerca de la veracidad del test y de su interpretación. En este artículo se evalúan la evolución de los tests de anticuerpos, la base de su especificidad y la validez de su interpretación. La aceptación de un test de anticuerpos del VIH como científicamente válido y fiable requiere de lo siguiente: (I) Una fuente de antígenos específicos del VIH; (II) Estandarización; (III) Determinación de la reproducibilidad del test. Una vez que se hayan cumplido estos criterios, y antes de la introducción de los tests de anticuerpos en la medicina clínica, debería determinarse la sensibilidad, especificidad y valores predictivos del test mediante el uso de un estándar oro, el VIH en sí mismo.
Proteínas consideradas como antígenos del VIH
Las proteínas que se considera que representan los antígenos del VIH se obtienen de cultivos estimulados mitogénicamente en donde los tejidos de los pacientes de sida son cocultivados con células procedentes de pacientes sin sida, normalmente líneas de células leucémicas consolidadas. Tras la detección de la enzima transcriptasa inversa (RT) en los cultivos, el sobrenadante, y con mas frecuencia los lisados celulares, se centrifugan en gradientes de densidad. Al material que se concentra a 1,16 g/ml se le considera que representa "VIH puro" y, por consiguiente, a las proteínas encontradas a esa densidad se las considera como antígenos del VIH. Se cree que las proteínas inmunogénicas del VIH están codificadas por tres genes, llamados gag, pol y env. El gen gag codifica al precursor p53/55, que entonces se parte en la p24/25 y la p17/18. El gen pol codifica la p31/32 y el gen env codifica al precursor p160, que se parte en la p120 y p41/p45 (1).
La proteína p120
La opinión generalmente aceptada es que la p120 y la p41 son productos de división de la p160, que se encuentra solamente en células infectadas y no en el virus. Sin embargo, la p120 solamente es un componente de los botones (salientes) en la superficie de las partículas de VIH; los botones se encuentran solamente en las partículas en gemación (inmaduras), y no en las partículas liberadas de la célula (maduras); las partículas inmaduras "se observan muy raramente" (2).
A pesar de estos hallazgos, cuando se testea el "VIH puro" contra los sueros de sida, aparecen bandas marcadas correspondientes a la p120 y la p160. La solución a estas contradicciones se encontró cuando se mostró que la p80 (vease más adelante) y "los componentes visualizados en la región de 120-160 kDa no se corresponden a la gp120 o su precursora, sino más bien representan oligómeros de la gp41" (3).
La proteína p41
La p41 es una de las proteínas detectadas tanto por el grupo de Gallo como el de Montagnier en los primerds aislamientos de VIH. Sin embargo, Montagnier y sus colegas observaron que los sueros de sida reaccionaban con una proteína p41 tanto en células infectadas con VIH como en células infectadas con HTLV-I, así como en células no infectadas, concluyendo que la banda p41 "podría deberse a contaminación del virus con actina celular, la cual estaba presente en los inmunoprecipitados de todos los fragmentos celulares" (4). Aunque el grupo de Gallo no encontró tal reacción con la p41 en células no infectadas, encontraron una proteína p80 y concluyeron que la reacción era "no específica" (5).
La actina es una proteína ubicua que se encuentra en todas las células, así como en las bacterias y algunos virus. Se ha demostrado que retrovirus bien conocidos, como el virus de tumor mamario de ratón y el virus de sarcoma de Rous, también contienen actina de origen celular, habiéndose propuesto que esta proteína juega un papel clave en el ensamblaje y la gemación retroviral (6,7). También se conoce que la oxidación de los grupos sulfhidrilo celulares, como es el caso de los pacientes de sida (8), tiene correlación con el ensamblaje de la actina polimerizada (9), y que el nivel de unión de anticuerpos de actina a las células está determinado por el estado fisiológico de las células. Por este motivo, se propuso esta unión de anticuerpos de actina a las células "como un marcador sensible para los linfoticos activados" (10).
Las plaquetas de individuos sanos también contienen una proteína p41-p45 que reacciona con los sueros procedentes de hombres homosexuales con sida y con púrpura trombocitopénica idiopática o inmune (ITP), y que "representa la unión no específica de IgG a la actina en la preparación de plaquetas" (11).
La proteína p32
En 1987 Henderson aisló la p30-32 y la p34-36 de "VIH purificado mediante doble banda" en gradientes de densidad de sacarosa. Comparando las secuencias de aminoácidos de estas proteínas con las proteínas DR de histocompatibilidad de Clase II, concluyeron que las "cadenas DR alfa y beta parecían ser idénticas a las proteínas p34-36 y p30-32 respectivamente" (12).
La proteína p24-p25
Actualmente, se cree que la detección de la p24 es sinónimo de aislamiento del VIH y de viremia. Sin embargo, aparte de una publicación conjunta con Montagnier, donde afirman que la p24 del VIH es singular, Gallo y sus colegas han declarado repetidamente que las p24 del HTLV-I y el VIH tienen reacciones cruzadas inmunológicas (13);
Genesca et al. (14) realizaron ensayos WB en 100 muestras negativas de ELISA procedentes de donantes de sangre sanos; se encontró que 20 tenían bandas que no satisfacían los criterios utilizados entonces (1989) por los bancos de sangre de un WB positivo. Se las consideró WB indeterminados (WBI), con la proteína p24 siendo la banda predominante en el 70% de los casos. Entre los receptores de sangre WBI, 36% fueron WBI después de 6 meses de la transfusión, pero asímismo lo fueron 42% de las personas que recibieron muestras WB negativas. Tanto los donantes como los receptores de sangre permanecieron sanos. Se concluyó que los resultados WBI "son demasiado comunes en donantes y receptores seleccionados aleatoriamente, y tales resultados no correlan con la presencia de VIH-1 o con la transmisión de VIH-1", "la mayoría de dichas reacciones representan resultados de falso positivo". Los anticuerpos a la p24 se han detectado en 1 de cada 150 individuos sanos, en 13% de pacientes sanos excepto por tener verrugas generalizadas, escogidos aleatoriamente, 24% de pacientes con linfoma de células T y pródomo , y en 41 % de pacientes con esclerosis múltiple (15).
El 97% de sueros de homosexuales con ITP y el 94% de sueros de homosexuales con linfadenopatía o sida contienen un anticuerpo que reacciona con un antígeno de membrana de 25 Kd encontrado en plaquetas procedentes de donantes de sanos y pacientes de sida, así como con un antígeno 25 Kd encontrado en células de riñón de mono verde, fibroblastos de piel humanos, y herpes simple cultivado en células de riñón de mono. Esta reacción no se produjo en sueros obtenidos de pacientes no homosexuales con ITP o púrpura trombocitopénica no inmune (11).
En cambio, el antígeno p24 no se encuentra en todos los pacientes VIH positivo, o incluso en pacientes de sida. En un estudio se utilizó la reacción en cadena de la polimerasa (PCR) y la p24 para detectar el VIH en pacientes en diversas etapas CDC, desde asintomáticos a sida. La p24 se detectó en el 24% de pacientes y ARN de VIH en el 50% (16).
En otro estudio, "En la mitad de los casos en que un individuo tenía un test p24 positivo, el individuo tuvo más tarde un test negativo sin tomar ninguna medicación que se esperase que afectara a los niveles del antígeno p24 ... el test es clínicamente errático y debería interpretarse con mucha cautela" (17).
La proteína p17/p18
Además de la banda p24, la banda p17/p18 es la banda más frecuentemente detectada en los WB de donantes de sangre sanos (18).
Los sueros de pacientes de sida se unen a una proteína p18 en células T infectadas con VIH estimuladas mitogénicamente, pero no a linfocitos no estimulados no infectados. Sin embargo, cuando los linfocitos son estimulados mitogénicamente, pero no infectados, los sueros de sida se unen a una proteína p18 en estos linfoticos no infectados (19). Un anticuerpo monoclonal (MCA) a la p18 del VIH, reacciona con células dendríticas en los tejidos linfáticos de diversos pacientes con ciertas enfermedades no relacionadas con el sida (20); y el "mismo patrón de reactividad estuvo presente en tejido normal obtenido de individuos no infectados como en el obtenido de individuos VIH positivo" (21).
Los pacientes de sida y aquellos en riesgo tienen niveles altos de anticuerpos a la proteína ubicua miosina (22), que tiene dos subunidades de pesos moleculares de 18.000 y 25.000. Vistos todos estos datos es difícil defender la visión de que las bandas p41 (y por tanto p160 y p120), p32, p24 o p18 representen proteínas específicas del VIH. Incluso si se pudiera mostrar que todas estas proteínas son específicas del VIH, no se puede asumir automáticamente que los anticuerpos que reaccionan con cada una de estas proteínas sean específicos a la infección de VIH.
Estandarización de los tests de anticuerpos del VIH
Un test de anticuerpos resulta significativo solamente cuando está estandarizado, es decir, cuando el resultado de un test concreto tiene el mismo significado en todos los pacientes, en todos los laboratorios y en todos los países. En las primeras reacciones antígeno-anticuerpo realizadas por los grupos de Montagnier (4) y Gallo (23) (fig. 1, 2) se encontró que: no todas las "proteínas del VIH" reaccionan con todos los sueros de los pacientes de sida, o incluso sueros de los mismos pacientes obtenidos en momentos distintos, y que sueros de pacientes de sida pueden reaccionar con proteínas distintas a las que se consideran como antígenos del VIH. Debido a éstas reacciones variables, un requitiso esencial fue el establecer criterios de lo que constituye un WB positivo.
Al principio, el grupo de Montagnier consideró la p24 como suficiente para definir un WB positivo, mientras que el grupo de Gallo consideró suficiente la p41. La mayoría, si no todos los laboratorios restantes, utilizaron los criterios recomendados por el CDC, concretamente la presencia de una banda p24 o p41. En 1987 fue obvio que esas bandas no eran específicas del VIH. Además, hasta 1987 "hubo tantos procedimientos WB como laboratorios realizando el ensayo" (24). Desde entonces, todos los principales laboratorios han cambiado sus criterios de interpretación del WB, pero en los Estados Unidos no hay todavía un criterio acordado nacionalmente, incluso entre los laboratorios principales:
En 1987 la Food and Drug Administration (FDA) autorizó un kit WB elaborado por DuPont. Este kit sigue siendo el único kit de WB con licencia y es utilizado por una minoría de laboratorios. Especifica unos criterios "extremadamente rigurosos" para un resultado positivo, concretamente "bandas específicas que representan tres productos génicos distintos: p24 (gag), p31 (pol), y una banda env, bien la gp41, la gp120 o la gp160" (24). La Cruz Roja Americana define un resultado positivo como la presencia de anticuerpos a, al menos, un producto génico de cada uno de los genes gag, pol y env, sin especificar que bandas.
La Association of State and Territorial Public Health Laboratory Directors/Department of Defence/CDC considera un WB positivo si dos de las p24, gp41 y gp120/160 son reactivas.
El Consortium for Retrovirus Serology Standardization (CRSS) define un WB positivo como la presencia de anticuerpos a al menos la p24 o p31/32, y la gp41 o gp120/160 (25). Todos los restantes laboratorios principales de los Estados Unidos que hacen el test del VIH tienen sus propios criterios. Para todos los laboratorios, un resultado negativo requiere la ausencia de todas las bandas, incluyendo bandas que no representan las "proteínas del VIH". Todos los demás patrones que no satisfacen los criterios de un test positivo o negativo de un determinado laboratorio se consieran como WBI por ese laboratorio.
De este modo, en la literatura científica no se ha publicado ningun estándar de bandas de un positivo WB. La Fig.0 está obtenida del manual de instrucciones de un fabricante de kit de WB, Bio-Rad. Aunque explica "Ejemplos de una muestra de suero de paciente reactiva típica, y la reacción con un control fuerte, débil y no reactivo", también se afirma que "Este ejemplo muestra solamente patrones de reactividad típicos, y no se debe utilizar como una referencia de comparación con resultados procedentes de muestras de suero desconocidas ... Las muestras de pacientes pueden mostrar diversos grados de reactividad con proteínas distintas, mostrando de este modo patrones de desarrollo de banda distintos ... Cada laboratorio que realiza el test del Western Blot debería establecer sus propios criterios para la interpretación de las bandas. De modo alternativo, la interpretación de las bandas puede dejarse al médico clínico".
Además de los problemas obvios asociados con la falta de estandarización, todas las interpretaciones anteriores llevan a problemas mayores:
Cuando se utilizan los criterios de la FDA para interpretar el WB, solamente un número mínimo (menos del 50%) de los pacientes de sida tienen un WB positivo, es decir, infectados con el VIH. Si se utilizan los criterios de la CRSS, el porcentaje de pacientes de sida que dan positivo aumenta al 79%
Más importante, incluso cuando se utilizan los criterios más estrictos, el 10% de las muestras de control, que incluyen "especímenes de centros e donantes de sangre", tienen un WB positivo (25).
Como ya se ha mencionado, Henderson y sus colegas mostraron que la p31/32 no es una proteína del VIH. Pinter y sus colegas mostraron que la p160 y la p120 son oligómeros de la gp41. También mostraron que el patrón de WB obtenido depende de muchos factores, incluyendo la temperatura y la concentración de dodecilsulfato sódico, utilizado para trastocar el "virus puro", concluyendo:
"La confusión en la identificación de estas bandas ha dado lugar a conclusiones incorrectas en estudios experimentales. De modo similar, algunos especímenes clínicos podrían haber sido identificados erróneamente como seropositivos, al suponer que estas bandas reflejaban reactividad específica contra dos componentes virales distintos, y cumplían un criterio de positividad verdadera o probable. La identificación correcta de estas bandas afectará a los estándares a establecer en la positividad del Western Blot: podría necesitar la reinterpretación de los resultados publicados" (26).
El hallazgo de que la banda p31/32 representa una proteína celular, y que la p120 y p160 son oligómeros de la p41, reduce los criterios de la CRSS y el de la Cruz Roja Americana a dos bandas, p24 y p41, que según el coronel Donald Burke "distan de ser específicas" (27). Los hallazgos anteriores reducen los criterios de la Association of State and Territory Public Health Laboratory Directors/Department of Defence/CDC a la p24 o la p41, generalmente aceptadas como que son no específicas.
A pesar de los datos anteriores, incluso en la actualidad se considera que las bandas p160, p120 y p41 representan distintas glicoproteínas de la envoltura viral. De hecho, las directivas actuales del la OMS consideran a un suero como positivo de anticuerpos al VIH-1 si "están presentes en la tira dos bandas de glicoproteína de envoltorio (con o sin) otras bandas específicas virales" (28).
Hasta la fecha, el sida en África se define en términos clínicos. Recientemente, el CDC recomendó la inclusión futura de pruebas serológicas de la infección por VIH en la definición africana del sida. El test recomendado es el ELISA (29), que no puede considerarse específico.
En Rusia, en 1990, de 20.000 tests de detección positivos "solamente se confirmaron 112" utilizando el WB como estándar oro, y en 1991, de aproximadamente 30.000 tests de detección positivos solamente se confirmaron 66 (30).
Según las definiciones de sida de Latinoamérica y el Caribe, los "signos clínicos de la infección por VIH" son confirmados "mediantes test de anticuerpos utilizando los métodos ELISA, inmunofluorescencia o WB". No se facilitan criterios para la interpretación del WB (31).
Reproducibilidad
Los problemas asociados con la reproducibilidad se pueden mostrar de la mejor manera con dos ejemplos. La fig.3 representa tiras WB de un espécimen de suero de un paciente con sida, testeado por 19 laboratorios que participaron en la segunda conferencia CRSS en estandarización del test WB (25). Como se puede ver, el patrón de banda obtenido con un mismo suero varía de laboratorio en laboratorio, aunque todos los laboratorios reportaron este espécimen como positivo.
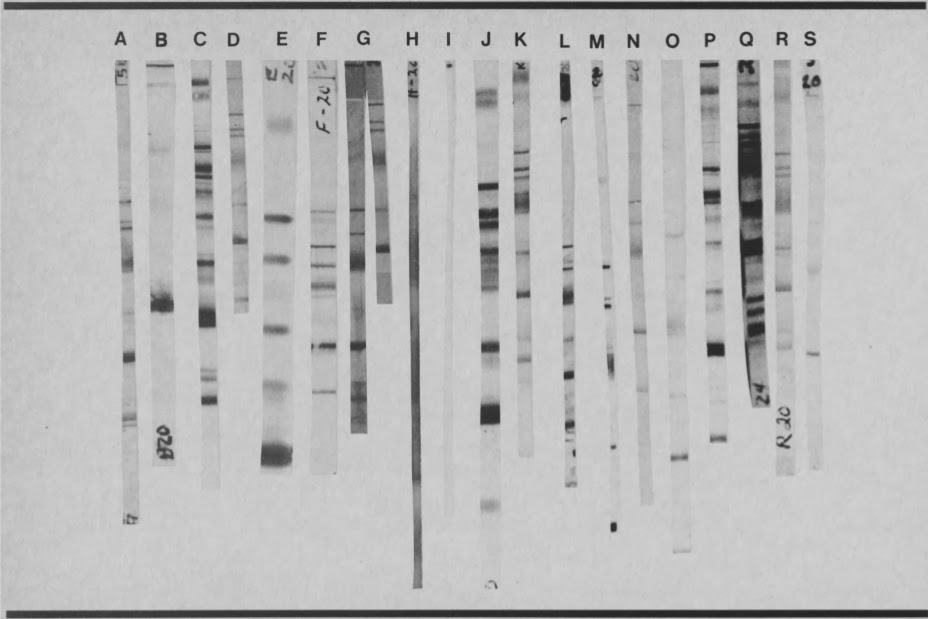
El Transfusion Safety Study (TSS) Group, en los Estados Unidos, envió aproximadamente 100 muestras de pacientes semanalmente a tres laboratorios de referencia para hacerles test de WB, en tres periodos separados de algunos meses. Junto a las muestras de los 100 pacientes, enviaron alícuotas procedentes de cuatro plasmas de control de calidad (QC), dos positivas y dos negativas. La positividad o negatividad al VIH "estuvo basada en la experiencia colectiva con cada plasma utilizando: (a) sistemas EIA con licencia de cinco fabricantes, (b) un ensayo de inmunofluorescencia, (c) IB en cuatro laboratorios de referencia, y (d) un ensayo de radioinmunoprecipitacion en un laboratorio adicional". (EIA = ELISA; IB = WB).
Las muestras fueron entonces enviadas a laboratorios de referencia, que eran conscientes del testeo de control de calidad, pero "las etiquetas y códigos no permitieron la identificación de los especímenes de QC como tales, o la vinculación con especímenes de QC previos.
QC1#(+) se envió 40 veces al laboratorio A, 5 veces al laboratorio B y 45 veces al laboratorio C. El laboratorio A informó de los siguientes patrones de bandas: p24, p32 y gp41/120, 7 veces; p24, gp41/120, 28 veces; p24 solamente, 5 veces. El laboratorio B reportó: p24, p32, gp41/120, 4 veces; p32, gp41/120, en una ocasión. El laboratorio C reportó: p24, p32, gp41/120, 26 veces; p24, gp41/120, 10 veces; p24, p32, dos; p24 solamente, 5 veces; "otros", una vez; sin bandas, una vez.
QC#2(+) se envió un total de 89 veces a tres laboratorios, y se reportó: p24, p32, gp41/120, 64 veces; p24, gp41/120, 19 veces; p24, p32, una vez; p32, p41/gp120, 4 veces; sin bandas, una vez.
Un total de 101 alícuotas de las dos muestras negativas de control de calidad, QC#3(-) y QC#4(-), fueron enviadas a los tres laboratorios. Estos reportaron: sin bandas, 67 veces; "otras" bandas, 13 veces; gp41 solamente, una vez; p24 solamente, 18 veces; p24, p32, gp41/120, dos veces.
Un panel especial de muestras de QC se envió a los laboratorios B, C, y a un laboratorio adicional D. El panel consistió en tres alícuotas de cada una de las ocho muestras, incluyendo los lotes QC#1(+), QC#2(+), QC#3(-) y QC#4(-).
En el análisis de los últimos resultados los autores afirmaron: "Solamente los reportes del Laboratorio C con el panel fueron consistentes con los datos acumulados de toda otra evaluación de reactividad ... El Laboratorio B reportó las tres alícuotas de QC#1 (+) como, respectivamente, positivas sobre la base de tres bandas (gp41, p55 y p65), indeterminadas sobre la base de una única banda (gp41), y negativas (no se observan bandas). Además, las las tres alícuotas de QC#6(-) se consideraron indeterminadas, porque solamente se vio una única banda (gp41). El Laboratorio D reportó una alícuota de QC#6(-) como positiva (p15, p24, p32, gp41, p65), y las otras dos alícuotas como negativas (no se observan bandas). También reportó una banda en p55 para las tres alícuotas de QC#3(-)"(32).
Al examinar estos resultados detallados, se debe tener en cuenta que se dieron en Laboratorios de Referencia, es decir, laboratorios de primera clase que constituyen solamente un pequeño número del total de laboratorios que realizan el test de WB en los Estados Unidos.
Además, muchos laboratorios continúan usando kits WB sin licencia debido al coste y los "criterios estrictos requeridos para interpretar el test autorizado" (33).
Especificidad de los tests de anticuerpos del VIH
La tarea de autentificar un test de diagnóstico nuevo en la medicina clínica requiere de un método alternativo independiente de determinar la presencia de la condición para la cual el test va a ser empleado. Este método, con frecuencia denominado estándar oro, es una condición sine qua non crucial, y representa el principio sobre el cual descansa la prueba de validez científica.
El único estándar oro posible para los tests de anticuerpos del VIH es el Virus de Inmunodeficiencia Humana en sí mismo. Claramente, el síndrome clínico y la disminución de células T4 no puede considerarse como un estándar oro. Aunque el VIH nunca se ha utilizado como estándar oro hay un consenso general respecto a que esta probada firmemente la especificidad de los tests de anticuerpos del VIH. Para el ELISA, las mejores cifras de Gallo, obtenidas a partir de pacientes de sida y 297 donantes de sangre sanos, fueron 97,7% de sensibilidad y 92,6% de especificidad, considerando los tests dudosos como positivos, y utilizando el síndrome clínico como estándar oro (34).
El Coronel Donald Burke y sus colegas del Walter Reed Army Institute de los Estados Unidos están acreditados por ser los que más a fondo han investigado el problema de definir la especificidad de los anticuerpos del VIH en una población grande, y se cree ampliamente que sus datos son los más actualizados y rigurosos. (35) Burke et al (36) testearon una subpoblación sana seleccionada rigurosamente de 135.187 individuos, elegidos según una muy baja prevalencia de infección de VIH, una décima parte de la un grupo mucho más amplio de candidatos (1,2 millones) para el servicio militar de los Estados Unidos.
A todos los candidatos se les hizo un test de detección con un ELISA inicial. Todos los tests ELISA reactivos fueron repetidos una vez. Entonces, se realizó un WB inicial y, si era diagnóstico o reactivo, se realizó un segundo WB en otro espécimen de sangre fresca. Inicialmente, los criterios de un WB positivo y diagnóstico fueron la "presencia de una banda en 41 kd, una combinación de las bandas 24 y 55 kd, o ambas.
A partir de mayo de 1987, se modifica el método de preparación de las tiras del blot, de modo que los anticuerpos a la gp120 y la gp160 podían detectarse de modo reproducible, y los criterios de un patrón de blot reactivo y diagnóstico se cambiaron a los de la Association of State and Territorial Public Health Laboratory Directors".
Se diagnóstico un positivo WB sí y solamente sí la primera y la segunda muestras de suero eran diagnósticas en el WB. Todas las muestras de WB diagnósticas fueron entonces analizadas con otros cuatro tests de anticuerpos. Se consideró un WB como "positivo verdadero si los cuatro ensayos en todas las muestras de suero disponibles procedentes de un candidato eran reactivas y diagnósticas", pero fue considerado un "falso positivo si los cuatro ensayos en todas las muestras de suero disponibles procedentes de un candidato eran no reactivas, no diagnósticas, o ambas".
De los 135.187 candidatos, hubo 16 tests positivos. En uno de estos, el suero no estuvo preparado para un testeo adicional, y un candidato se negó a proporcionar una segunda muestra. El suero procedente de 27 de las 29 muestras de los 15 candidatos encontrados positivo fue testeado mediante los otros cuatro tests de anticuerpos. Catorce muestras fueron positivas a los cuatro ensayos, y los cuatro fueron negativas para un candidato.
De aquí Burke y sus colegas calcularon que la tasa de falsos positivos es de 1 cada 135.187 o 0,0007%. También especularon en las implicaciones que este dato tendría en la población entera de 1,2 millones de candidados. Calcularon la prevalencia total en 1,48 por 1.000 en el grupo entero, equivalente a 200 para 135.187. Asumiendo que la tasa de falsos positivos es la misma para la población completa, estimaron que puesto que habrá 200 tests positivos para 135.187 personas, de las cuales solamente una será un falso positivo, entonces el "valor predictivo de un diagnóstico positivo en el programa es de 99,5%, y una especificidad de 99,9% (35,36).
Se puede criticar bastante el razonamiento de Burke y sus colegas:
(I) No existe un estándar oro para definir la infección de VIH. El testear los positivos WB en los 15 candidatos restantes contra otros cuatro tests de anticuerpos no posibilita un establecimiento independiente de infección por VIH "verdadera", puesto que son el mismo test;
(II) Ellos definen: (a) los tests positivos verdaderos como muestras que tienen test positivo repetidamente con cuatro tests similares. (b) los tests falsos positivo como muestras que tienen un test negativo repetidamente con cuatro tests similares. El número de muestras testeadas y las repeticiones se definen arbitrariamente. Sería imposible decir cual sería el resultado si, por ejemplo, se repitieran los tests ELISA tres veces en vez de dos, o si las muestras que dieron test negativo en el primer ELISA fuesen testeadas de nuevo con otro ELISA o WB. Hay informes bien documentados en los que el ELISA es negativo y el WB positivo (37). (c) la tasa de falsos positivos como el número de resultados falso positivo dividido por el número de muestras testeadas. Estas definiciones no se parecen en absoluto a las descritas en textos estándar (38). Las definiciones correctas son: (i) Un positivo verdadero es un test positivo que ocurre en un individuo que está infectado por VIH, según la definición mediante un estándar oro independiente; (ii) Un falso positivo es un test positivo que ocurre en un individuo que, mediante la aplicación del estándar oro, no tiene infección por VIH (pero no está sano necesariamente); (iii) la tasa de falsos positivos es el número de tests falsos positivos dividido entre todos los tests positivos, tanto negativos como positivos.
(III) Las premisas de Burke et al son totalmente contrarias a las de Gallo et al, donde todos los resultados de test positivo en individuos sanos se consideran como falsos positivos. Basandose en las premisas de Gallo y sus colegas se debe considerar a todos los dieciséis casos como falsos positivos, ya que no hay una razón convincente para considerar a los candidatos militares sanos significativamente diferentes de los donantes de sangre sanos.
(IV) La extrapolación de Burke al total de 1,2 millones de candidatos no es válida. Esta extrapolación solamente puede realizarse si los 135.187 candidatos se seleccionasen aleatoriamente del grupo entero, lo cual no se hizo. En el resto de la población la tasa de falsos positivos podría haber sido mucho mayor, por ejemplo, como resultado de altas concentraciones de globulinas en general, y de autoanticuerpos en particular.
Es imposible obtener el valor predictor positivo de 99,5% que indican sin tener conocimiento de la sensitividad del test WB y la prevalencia de la infección de VIH verdadera (38), incluso si la especificidad y la extrapolación fuesen correctas.
(V) Es imposible definir la especificidad, la sensiblidad y el valor predictivo con el algoritmo utilizado por Burke y sus colegas. Lo mejor que pueden hacer con su algoritmo es determinar la reproducibilidad del ELISA y el WB. En este sentido, en el estudio más grande de Burke de 1,2 millones de candidatos militares sanos, aproximadamente el 1% de todos los iniciales, 50% de todos los ELISA's repetidos fueron positivos; y 30-40% de los primeros WB fueron positivos y 96% de los segundos WB fueron positivos. Es decir, el estudio más grande de Burke muestra: (a) 6.000 individuos con un ELISA positivo inicial, pero posteriormente uno negativo. (b) 4.000 individuos con dos ELISA's positivos, seguido por un WB negativo. (c) 80 individuos con dos ELISA's positivo, un WB inicialmente positivo y un WB repetido negativo.
A esto no se le puede considerar un problema trivial, ya que: (I) tanto el ELISA como el WB están considerados como altamente sensitivos y específicos. (24) (II) Algunos miles de individuos sanos tienen anticuerpos que reaccionan con las "proteínas del VIH", pero que no son finalmente considerados como infectados con el VIH; (III) Incluso en los mejores laboratorios, 80 de los candidatos sanos de Burke serían diagnosticados como infectados por VIH ya que, a diferencia de Burke, solamente se realiza un WB.
El problema llega a ser más serio cuando uno se da cuenta de que en septiembre de 1987, fecha en la cual, basándose en los tests de anticuerpos, se aceptó por lo general una relación causal entre el VIH y el sida, entonces para un positivo ELISA o un positivo WB, una banda (bien la p24 o la p41) fue suficiente para confirmar la infección por VIH.
En la actualidad, la opinión general es que los tests ELISA tienen una "sensibilidad y especificidad de sobre el 98%, muchos aproximándose al 100%" (24), y la definición del sida del CDC "acepta un test de detección reactivo de anticuerpos de VIH sin una confirmación por un test adicional, porque un resultado repetidamente reactivo de test de detección , en combinación con un enfermedad indicadora, es altamente indicativa de una enfermedad de VIH verdadera" (39). (test de detección = ELISA).
Burke et al, al igual que Gallo et al, determinó la especificidad sin hacer referencia a los individuos enfermos. La definición de especificidad requiere que el test sea evaluado en personas que no tienen la enfermedad que está bajo examen, incluyendo individuos enfermos que tienen otras enfermedades donde los anticuerpos, algunos de los cuales pueden interactuar con los antígenos del VIH, pueden producirse por otras causas. La especificidad de los tests de anticuerpos del VIH debe determinarse mediante el testeo de individuos que están inmunodeprimidos y/o tienen síntomas y señales clínicas similares al sida, pero que no se considera que tienen sida o infección con VIH. Este punto se ilustra muy bien con los tests serológicos de la sífilis. Una persona sana que no está infectada con el Treponema pallidum daría un test positivo (falso positivo) muy raramente.
Sin embargo, algunos autores atestiguan la presencia, en varios desordenes no relacionados, de tests biológicos falsos positivos a la sífilis (BFPS), que pueden ocurrir en pacientes con anemia hemolítica autoinmune, lupus eritematoso sistémico (SLE), púrpura trombocitopénica idiopática (inmune), y lepra, así como en adictos a drogas. Más de un 20% de los adictos a drogas tienen un test positivo y tienen la mas alta incidencia de BFPS's (40).
También se vio que las personas con BFPS "tienen una alta frecuencia de otras anormalidades serológicas, incluyendo factores antinucleares, autoanticuerpos, y alteraciones de la gamma globulina". Esto llevó a los investigadores a concluir que "una reacción BFP es con frecuencia un marcador de un desorden no identificado del sistema inmune que predispone a las enfermedades autoinmunes" (40). Es significativo que en una alta proporción (14%) de pacientes de sida se encontro también que tenían una serología de sífilis de falso positivo (41).
Al menos dos grupos de investigadores plantearon la posibilidad de que un test de anticuerpos al VIH en africanos y usuarios de drogas intravenosas pueda deberse a una reacción BFP. Jaffe et al (42) testearon 1129 muestras de suero procedentes de usuarios de drogas intravenosas y 89 controles de usuarios normales. Se recogieron todas las muestras durante 1971-1972 y se testearon por dos ELISA's y un WB comerciales. Diecisiete de las muestras de los usuarios de drogas intravenosas fueron positivas, y ninguna de la de los controles.
Concluyeron: "Según nuestros datos de Western Blot positivos, parece que los usuarios de drogas parenterales pueden haberse expuesto al HTLV-III o a un virus relacionado tan pronto como en 1971. Una explicación alternativa pero igualmente viable es que la seropositividad HTLV-III detectada en estos espécimenes represente falsos positivos o reacciones no especificas".
Biggar y sus colegas (43) hallaron que en africanos sanos la probabilidad de encontrar un test de anticuerpos del VIH positivo se incrementaba significativamente con el aumento de los niveles de complejo inmune. Concluyeron que "la reactividad en el análisis ELISA y WB podría ser no específica en los africanos ... la causa de la no especificidad necesita clarificarse para poder determinar como podrían afectar a la seroepidemiología de retrovirus en otras áreas distintas a África, como el Caribe y Japón.
El que un positivo WB en todos los individuos puede representar una reacción BFP se sugiere por datos procedentes de la retrovirología en general, y de la relización de tests de anticuerpos del VIH en particular.
Se sabe que todos los anticuerpos, incluyendo los MCA, son poliespecíficos y son capaces de reaccionar con antígenos inmunizantes, así como con otros componentes propios y no propios (44,45). En relación con los retrovirus, en la literatura científica abundan datos que muestran de modo convincente la presencia extendida de interacciones no específicas entre antígenos retrovirales y anticuerpos no relacionados. Mucho de este trabajo ha aparecido como resultado de la búsqueda de un origen viral para las neoplasias animales y humanas (46-50).
En 1975 Gallo descubrió que los pacientes con leucemia tienen una infección generalizada a un retrovirus que Gallo sostuvo haber aislado a partir de cultivos y tejidos frescos de estos pacientes, y que llamó HL23V. Gallo sugirió que este virus estaba etiológicamente asociado con la enfermedad, pero posteriormente se mostró que el HL23V era un "cóctel" de dos virus de mono.
En 1980 Gallo descubrió el HTLV-I, que él y sus colegas sostuvieron que causaba leucemia de células T adultas. Hasta el 25% de los pacientes de sida tienen anticuerpos a este virus (51), pero los pacientes de sida no desarrollan leucemia con más frecuencia que la población general. Esto solamente puede interpretarse como que el HTLV-I no causa leucemia de células T adultas, o que algunos anticuerpos retrovirales detectados en pacientes de sida no son específicos.
En 1986 Essex obtuvo datos serológicos y de aislamiento de otro "retrovirus humano", el HTLV-IV. Se mostró posteriormente que el HTLV-IV de Essex era un virus de mono, ahora llamado Virus de Inmunodeficiencia Simio.
Que un WB positivo puede no ser una prueba de infección por VIH , sino que es solamente un marcador no específico de sida, se sugiere de los siguientes datos:
En los drogadictos, hay una fuerte asociación entre altos niveles de globulina en suero y un test de anticuerpos de VIH positivo, siendo ésta la "única variable que continuó siendo significativa en un modelo de regresión logística" (52). En niños, utilizando el WB como estándar oro, la hiperglobulinemia identificó a los niños infectados con VIH con una especificidad del 97% (53). Se hizo el test de anticuerpos del VIH a sesenta y tres sueros obtenidos de 23 pacientes antes e inmediatamente después de una infusión de inmunoglobulina, utilizando el WB. De los 63 sueros, 52 (83%) fueron positivos. "Algunas muestras testeadas en un radioinmunoensayo de p24 de HTLV-III también fueron positivas. La cantidad de anticuerpos detectada fue máxima inmediatamente después de la infusión, y disiminuyó entre infusiones" (54).
A un individuo se le dieron seis inyecciones de 5 ml de suero de un donante Rh+, administradas en intervalos de 4 días. "Se mostró que el suero del donante era negativo al ELISA de anticuerpos y antígenos del VIH, de modo que se tomó sangre de su mujer y su hijo". "Se mostró que la sangre tomada tras la primera inmunización era negativa al ELISA y al ensayo de inmnunoblot de anticuerpos del VIH . Tras la segunda inmunización, se monitorizó una señal débil en el ELISA, ligeramente por encima del nivel de corte. Tras la tercera inmunización, la señal fue más fuerte y el inmunoblot reveló una interacción perceptible con las proteínas p17 y p55. Se monitorizó una señal aún más fuerte tras la quinta inmunización. La interacción con la p17, la p31, la gp41 la p55 y otras proteínas era evidente" (55).
Puesto que los individuos de los grupos principales de riesgo el sida, es decir, los hombres gay, los usuarios de drogas y los hemofílicos están expuestos a muchas sustancias externas, como el semen, las drogas, el factor VIII, sangre y componentes sanguíneos; y los individuos que pertenecen a los grupos anteriores desarrollan habitualmente infecciones no relacionadas con el VIH, se esperaría que estos individuos tuviesen altos niveles de anticuerpos dirigidos contra otros antígenos distintos a los del VIH. De hecho, en la actualidad, existen datos de que los individuos con sida, complejo relacionado con el sida (ARC) y aquellos en riesgo tienen complejos inmunes circulantes y factor reumatoide, así como anticuerpos anticardiolipina, anti factor nuclear, anticelular, antiplaquetas, anti globulos rojos, antiactina, antiADN, antitubulina, antitiroglobulina, antialbúmina, antimiosina, antitrinitrofenil y antitimosina (22,56).
Se han encontrado autoanticuerpos antilinfocito en 87% de los pacientes VIH positivo, y sus niveles correlan con el estado clínico. (57,58). A diferencia de los sueros normales, se halló que el 37% de los sueros VIH positivos eran positivos a retrovirus Tipo D (59), mientras que se piensa que el VIH es un Lentivirus.
También se conoce que los niveles IgG de suero son más altos en donantes de sangre negros que en caucásicos (60); que algunos grupos de riesgo, consumidores de drogas y hombres homosexuales están expuestos a altos niveles de agentes mitogénicos, semen y nitratos (61,62), y que los animales tratados con tales agentes desarrollan anticuerpos que reaccionan con antígenos retrovirales (63).
Que un test de anticuerpos VIH positivo podría ser el resultado de una estimulación antigénica, distinta al VIH, se sostiene adicionalmente con los siguientes datos:
(I). Se cree que el VIH se transmite mediante jeringuillas infectadas, sin embargo tuvieron un test positivo más prostitutas que usan drogas orales (84%), que las que usan drogas intravenosas (46%) (64);
(II) "Los ratones de las cepas autoinmunes MRL-lpr/lpr y MRL-+/+ produjeron anticuerpos contra la gp120". Se mostró que los ratones que habían sido expuestos a linfocitos T procedentes de otra cepa murina producían anticuerpos contra la gp120 y la p24 del VIH (65).
(III) Los receptores de sangre negativa seroconvierten y desarrollan sida, mientras que los donantes permanecen sanos y seronegativos (66).
(IV) En individuos sanos, parejas de individuos positivos al VIH, receptores de transplantes de órganos y pacientes con SLE, un WB positivo puede revertir a negativo cuando cesa la exposición al semen, se administra la terapia inmunosupresiva o hay una mejora clínica (67,68,69);
(V) Mientras la frecuencia de los tests de anticuerpos VIH positivo en donantes de sangre sanos y candidatos militares es baja, los pacientes de tuberculosis (TB), incluyendo aquellos con TB localizada en los pulmones, tanto en los Estados Unidos (70) como en África (71), tienen una alta frecuencia de hasta del 50% de WB's positivos. En los Estados Unidos (72) (26 hospitales estudiados), los pacientes que no están en riesgo de desarrollar sida y que no tienen enfermedades infecciosas, tienen una alta tasa de WB positivos (1,3% a 7,8%).
Los datos anteriores podrían interpretarse como prueba de que el VIH se está extendiendo entre la población heterosexual, o bien que los tests de anticuerpos del VIH no son específicos. Que es en verdad este último caso se sugiere por el hecho de que en 1988, en los Estados Unidos (73), se informó que solamente aproximadamente 66 hombres blancos habían tenido "sida adquirido de modo heterosexual". En 1992, en Nueva York, se informó que solamente 11 hombres tenían sida debido a una infección heterosexual (74).
Rodríguez y sus colegas (75) hallaron que los indios del amazonas que no tienen contacto con individuos fuera de sus tribus y no tienen sida tienen un 3,3 - 13,3% de tasa de seropositividad del WB al VIH, dependiendo de la tribu estudiada.
En otro estudio (76), se vio que el 25% - 41% de los pacientes de malaria venezolana tenían un WB positivo, pero no tenían sida. Los datos anteriores significan que bien el VIH no causa sida "incluso en la presencia de graves trastornos inmunoreguladores característicos de la malaria aguda", como Rodriguez et al concluyeron, o bien que los tests de anticuerpos del VIH no son específicos.
Los problemas asociados con la especificidad del WB podrían evitarse con el uso del único estándar oro adecuado, el aislamiento del VIH. Hasta la fecha esto no se ha realizado, y basándose en los problemas asociados con el aislamiento del VIH, podría no ser nunca factible.
Aislamiento del VIH
Es obvio que el aislamiento de un virus puede utilizarse como estándar oro solamente si proporciona datos genéticos, virológicos y moleculares concluyentes de la existencia de un virus singular. Con los retrovirus, como primer paso hacia este objetivo, uno debe encontrar partículas con características morfológicas similares a otros retrovirus, y demostrar que estas partículas tienen un conjunto único de componentes estructurales, incluyendo que el ARN y las proteínas pertenezcan solamente a estas partículas, y no a otra entidad.
Peyton Rous (77) está reconocido como el que descubrió y aisló el primer retrovirus. En 1911 fue capaz de inducir tumores repetidamente en una determinada raza de pollos por medio de filtrados libres de células derivados de tumores.
Rous consideró que bien un "organismo parasitario pequeño" o un "estimulante químico" podría ser la base de sus observaciones; no obstante, los filtrados inductores de tumor llegaron a ser conocidos como "virus filtrables" u oncovirus.
En 1950, en cultivos de animales y en tejidos frescos, especialmente tejidos de tumor, a las partículas que más tarde se atribuyeron a los retrovirus se las pudo detectar sin dificultad con microscopía electrónica (ME).
En 1970, se descubrió en oncovirus la enzima transcriptasa inversa (RT), que transcribe ARN en ADN. Debido a esto, en los años 70 a los oncovirus se los conoció como retrovirus.
En la década precedente, se introdujo la centrifugación por gradientes de densidad para separar y aislar partículas subcelulares, incluyendo a los virus.
Se vio que algunos constituyentes celulares tenían la misma densidad de flotación que los virus, debido a lo cual cuando los virus eran aislados a partir de los cultivos celulares, los mejores resultados podían obtenerse con fluidos de sobrenadante, que tenían una alta concentración viral y pocos contaminantes celulares.
Esto fue más satisfactorio para virus no citopáticos y mediante condiciones de cultivo que mantenían una viabilidad celular máxima. La mayoría de los retrovirus animales (las excepciones se llaman virus de inmunodeficiencia animal) satisfacen las condiciones anteriores.
Aprovechando las propiedades retrovirales anteriores, se puede obtener a 1,16 g/ml, mediante suspensión y sedimentación repetida en gradientes de densidad de sacarosa, una concentración relativamente pura de partículas retrovirales, es decir, obtener partículas retrovirales separadas de todo lo demás, y de este modo aislarlas (78).
Sin embargo, como muchos retrovirólogos importantes señalan, podría no poder evitarse la contaminación de la preparación viral con partículas con aspecto de virus, que contienen RT, pero que podría no ser nada más que "fragmentos celulares", microsomas de células rotas, "vesículas membranosas que pueden contener otros constituyentes celulares, incluyendo ácidos nucléicos", especialmente cuando se induce una "lisis inadvertida de células". (79,80,81).
Debido a esto, para probar que el material que se concentra a 1,16 g/ml contiene nada más que partículas "sin diferencias aparentes en las apariencias físicas", y que las partículas eran realmente retrovirus, cada preparación de retrovirus se analizó adicionalmente utilizando los siguientes ensayos: (1) Físico: microscopía electrónica (ME) para el recuento del virus, su morfología y su pureza; (2) Bioquímico: actividad de RT, ARN viral y celular, análisis de gel de proteínas completo de las proteínas virales y del huesped, y de los ácidos nucléicos. (3) Biológico: infectividad in vivo e in vitro (78,82).
A diferencia de los cultivos de virus de animales, donde la concentración de partículas es muy alta (104-105 unidades infecciosas por mililitro), en los cultivos-cocultivos de sida la concentración de partículas es baja, tan baja que tanto el grupo de Gallo como el de Montagnier tuvieron dificultades en dectectarlas.
A diferencia de la mayoría de los retrovirus animales, al VIH se le considera como un virus citopático. Si esto es así, entonces los sobrenadantes de cultivo celular contrendrán muchos constituyentes celulares. Si, como se ha propuesto recientemente, "un mecanismo único singular", la apoptosis producida por el VIH , puede explicar la muerte de células T4 (83), entonces el sobrenadante también debe contener cuerpos apoptóticos, es decir, fragmentos celulares ligados a la membrana que (como muchos retrovirus) geman de la superficie de la célula.
Puesto que el tamaño y composición (algunos contienen cromatina picnótica) de los cuerpos apoptóticos varían ampliamente (84), sería de esperar que algunos de estos fragmentos se concentren también a 1,16 g/ml.
Es significativo que los cultivos-cocultivos de sida no tienen una viabilidad máxima, y la mayoría, si no todos, de los supuestos "aislamientos del VIH" han sido a partir de lisados celulares. Además, y lo más importante, tras una búsqueda exhaustiva en la literatura del sida no se encontraron micrografías electrónicas del material que se concentra a 1,16 g/ml; todas las micrografías electrónicas son de partículas encontradas en los cultivos celulares.
De este modo, es imposible saber si el material: lípidos, proteínas y ácidos nucléicos, que se concentra a 1,16 g/ml (las "partículas de VIH puras"), contiene alguna de esas partículas en absoluto, y si tales partículas estuvieran presentes, cual es su pureza.
Los datos disponibles actualmente indican que solamente el 20% de las proteínas que se concentran a 1,16 g/ml son "proteínas del VIH", el resto son celulares, incluyendo las proteínas beta 2 microglobulina y HLA-DR (4,4%) (12,85).
Por tanto, incluso si hay partículas a 1,16 g/ml y todas las proteínas que se supone que son VIH están incorporadas en la partícula de VIH, el material que se concentra a 1,16 g/ml no puede considerarse como "VIH puro".
Al contrario, "Gran parte de la proteína viral secretada de las células infectadas por VIH no es de partículas, y la proporción de (por ejemplo) p24 en viriones es una función del genotipo viral y la antigüedad del cultivo. En casos extremos, menos del uno por ciento del total de p24 y gp120 presente [en el cultivo] lo está en viriones" (86). De hecho, la p24 se libera de "células infectadas independientemente de las partículas de virus infecciosas" y la RT (87,88). Debe señalarse que los términos de la literatura del sida: "VIH", "aislamiento del VIH", "partículas puras", "partículas de virus", "viriones" y "partículas infecciosas", tienen una variedad de significados incluyendo todos los siguientes, pero en su mayor frecuencia sin prueba de la presencia de ninguna partícula: (a) "ARN envuelto en proteína" (89); (b) material procedente de los sobrenadantes de cultivo celular que pasa a través de filtros celulares ajustados, pero a través del cual podrían pasar organismos tales como los micoplasmas (90);(c) el sedimento obtenido mediante ultracentrifugación simple del sobrenadante del cultivo (91); (d) recientemente, con mucha frecuencia, detección en los cultivos de sida de la p24 (92,93).
En el primer reporte de "aislamiento del VIH", el grupo de Montagnier detectó una "actividad de transcriptasa inversa" "temporal" en un cultivo estimulado mitogénicamente procedente de las biopsias de ganglio linfático de un hombre gay con linfadenopatía. En linfocitos de cordón umbilical estimulados mitogénicamente cultivados con sobrenadante procedente de los cultivos anteriores, reportaron partículas con aspecto retroviral (RVP) tipo C en los cultivos, RT, y antígenos que reaccionaban con sueros pre-sida, en el material que se concentraba a 1,16 g/ml (94).
El grupo de Gallo no consideró que la detección de lo anterior representase un "aislamiento verdadero", "... el virus no se ha transmitido a una línea de células de crecimiento permanente para un verdadero aislamiento, y por tanto ha sido dificil de obtener en cantidad" (94).
Sin embargo, aunque el grupo de Gallo utilizó una línea de celulas permanente para el "aislamiento de VIH", no informaron de nada más que de los mismos fenómenos que el grupo de Montagnier.
No obstante, en la actualidad, se considera que la detección de los fenómenos anteriores constituyen un "verdadero aislamiento", y su detección en un cultivo similar se considera como prueba de infectividad. Sin embargo, el aislamiento se define como separar el virus de todo lo demás, y no la detección de algunos fenómenos atribuidos al virus (RT, reacciones antígeno-anticuerpo [WB]), o similares a él (partículas).
Ciertos fenómenos pueden utilizarse para detección viral sí y solamente sí, los fenómenos se han identificado como específicos del virus, mediante la utilización del virus aislado como estándar oro.
Aunque esto no se ha realizado, los datos indirectos disponibles actualmente (es decir, los datos que se han obtenido sin un estándar oro) procedentes de la retrovirología general y la investigación del sida, indican que la RT, las RVP y las reacciones antígeno-anticuerpo no son específicas del VIH (o incluso de los retrovirus).
La especificidad de las reacciones antígeno-anticuerpo ya ha sido analizada y no se mencionará más. En cualquier caso, esta reacción no puede utilizarse como un estándar oro para el WB, ya que un test no puede ser su propio estándar oro.
Transcriptasa inversa
En toda la investigación del VIH, se considera prueba de actividad de RT del VIH a la copia del iniciador-patrón An.dT15 cuando se incuba con el sobrenadante del material que se concentra a 1,16 g/ml, a partir de los cultivos-cocultivos de sida. En muchos casos, esta actividad se considera como sinónimo de "aislamiento del VIH", y se utiliza para cuantificar el virus.
Sin embargo: (a) También se copia el mismo iniciador-patrón cuando se incuba con material que se concentra a 1,16 g/ml procedente de cultivos de células T leucémicas (95) y espermatozoides normales no infectados (96). Tanto el An.dT15 como el Cn.dG15 son copiados por material que se concentra a 1,16 g/ml originado a partir de linfocitos no infectados normales pero mitogénicamente estimulados (95,97). (b) El An.dT15 se copia no solamente mediante la RT, sino también mediante dos (beta y gama) de las tres polimerasas de ADN celulares. De hecho, en 1975, una Conferencia Internacional de polimerasas de ADN eucarióticas definió la polimerasa de ADN gama como la enzima celular que "copia An.dT15 con alta eficiencia pero no copia bien el ADN" (98). De este modo, la copia del iniciador-patrón An.dT15 no puede considerarse sinónimo de la presencia de RT de VIH.
Detección de partículas
Los retrovirus son partículas infecciosas con una envoltura de unos 100 a 120 nm de diámetro, con un núcleo compuesto de una cubierta de proteínas y un complejo de ribonucleoproteína. Los retrovirus se clasifican en tres subfamilias: Spumavirinae, Lentivirinae y Oncovirinae. Los retrovirus que pertenecen a esta última se dividen en partículas de Tipo A, B, C y D.
No obstante, algunos de los mejores retrovirólogos no consideran el encontrar "partículas con aspecto viral que se parecen morfológicamente y bioquímicamente" a los retrovirus como prueba de la existencia de tales virus (99).
En los años 70, se observaron con frecuencia tales partículas en tejidos leucémicos humanos (99), cultivos de tejidos embrionarios (100,101), y "en la mayoría, si no todas, las placentas humanas" (102). Sin embargo, continúan siendo "un problema misterioso e importante que queda por resolver" (103).
Todos los investigadores del sida consideran a las partículas detectadas en cultivos-cocultivos de sida como VIH. Sin embargo:
(I) No hay acuerdo acerca de a qué Género o incluso Subfamilia de retrovirus pertenecen. Algunas veces no hay acuerdo incluso dentro del mismo grupo. Por ejemplo, el grupo de Montagnier informó inicialmente del VIH como un oncovirus Tipo C (4), posteriormente como un oncovirus Tipo D (104), y posteriormente como perteneciente a una Subfamilia distinta de retrovirus, la Lentivirinae (105). Sin embargo, las "partículas de VIH" en monocitos difieren de los Oncovirus Tipo C y de los Lentivirus (106).

(II) A pesar de lo anterior, Gelderblom et al presentaron un modelo de VIH (fig 4) que tiene una morfología y composición bien definidas, incluyendo protuberancias de superficie hechas de p120, una proteína que se considera que juega un papel crucial en la citopatogénesis, y que es indispensable para la infectividad del VIH (107). El modelo ha sido aceptado y es bien conocido. Sin embargo, el mismo grupo, utilizando ME y microscopía electrónica inmune han mostrado que: (a) las protuberancias solamente se encuentran en partículas inmaduras (en gemación). Las partículas inmaduras se "observan muy raramente" y solamente se ven "en células dañadas metabólicamente" (2,108); (b) las partículas maduras son "apenas etiquetadas, si no para nada" por los sueros de sida y ARC. Las partículas inmaduras son "altamente etiquetadas", pero también lo es el resto de la célula de donde están gemando, que "puede deberse al hecho de que los sueros inmunes naturales son ciertamente poliespecíficos" (2,109); (c) al igual que los sueros, los anticuerpos a la p120 reaccionan preferentemente con partículas inmaduras (107). Los MCA contra las proteínas gag etiquetan las partículas maduras, pero también etiquetan las partículas VIH-2 y las partículas del virus de inmunodeficiencia humana (110); (d) en las partículas de VIH, incluyendo su membrana, Gelderblom et al (111), así como otros (112), detectaron muchas proteínas celulares, pero con la posible excepción de los "cuerpos laterales", estas proteínas no están incluidas en el modelo idealizado del VIH. (III) Los "cultivos infectados por VIH" de células T y monocitos contienen, además de partículas con las morfologías atribuidas al VIH, muchas otras "partículas virales" distintas de cualquiera de las "partículas de VIH" (106, 111, 113, 114). Las células H9 no infectadas por el VIH, a partir de las cuales tienen su origen la mayoría de las ME publicadas, así como otras células utilizadas para el "aislamiento del VIH", CEM, C8166, células B transformadas en EBV, y linfocitos de sangre de cordón, expresan partículas de aspecto viral en gemación, si bien son algo diferentes de las partículas aceptadas como VIH (115). Los datos anteriores llevan a preguntas no solamente respecto al origen y papel de las "partículas que no son de VIH", sino también al de las "partículas de VIH", y en cuanto a qué partículas, si alguna, se concentran a 1,16 g/ml. (IV) Las partículas de tipo C en gemación y las maduras aparecen en células de linfoma dañadas metabólicamente pero no infectadas con el VIH (116). Se han encontrado "partículas retrovirales"relacionadas antigénicamente con el VIH en cultivos de fragmentos de glándulas salivales procedentes de pacientes con el síndrome de Sjorgen (117).
El hallazgo independiente de partículas "con aspecto viral" en los glanglios linfáticos de pacientes de sida con linfadenopatía (118), y de proteínas en los ganglios linfáticos que reaccionaban con MCA a la p55, p24 y p18 (119) se interpretaron como prueba de que las "partículas con aspecto viral" eran VIH. Sin embargo:
(I) Los MCA a la p18 reaccionan con tejidos linfáticos de pacientes que sufren un número de enfermedades no relacionadas con el sida, y también de individuos sanos (20,21);
(II) Al igual que en cultivos-cocultivos de sida, en los ganglios linfáticos de pacientes con sida y linfadenopatía generalizada persistente, además de las "partículas de VIH" se encuentran también partículas distintas a las de VIH (120).
(III) Más importante, en el único estudio de ME (121) in vivo o in vitro en donde se utilizaron controles adecuados, y en donde se realizó un exhaustivo examen a ciegas del material de test y de control, se encontraron partículas de virus indistinguibles del VIH en diversas linfadenopatías reactivas no asociadas al VIH, llevando a los autores a concluir: "La presencia de tales partículas no indican, por sí mismas, infección con el VIH".
Comentarios acerca del "aislamiento"
Se puede concluir entonces que ni la reacción de antígeno-anticuerpo, ni las partículas ni la RT pueden considerarse como específicas de los retrovirus. Incluso si lo fuesen, su hallazgo no puede considerarse como sinónimo de detección de un retrovirus adquirido externamente, como se sostiene en el caso del VIH. Tales hallazgos podrían representar la expresión de retrovirus endógenos (vide infra) u otros retrovirus exógenos. Últimamente "algunos laboratorios reportaron actividad retroviral [RT, partículas] en células de pacientes que parece no estar infectados con el VIH", una actividad que se dice que es "de retrovirus endógeno" (122).
La línea celular utilizada con mayor frecuencia en la investigación del sida es la línea de células leucémicas H9. Se sabe que la H9 es un clon de la HUT78, que se deriva de un paciente con leucemia de células T adultas. Puesto que se acepta que el agente causante de esta leucemia es el HTLV-I, otro retrovirus exógeno, los cultivos de H9 deberían tener tanto RT como partículas retrovirales incluso en la ausencia del VIH.
Puesto que el 25% de los pacientes de sida tienen anticuerpos al HTLV-I, sobre el 25% de los cultivos deberían tener, además de partículas y RT, un WB positivo aL HTLV-I. Sin embargo, puesto que las proteínas del VIH y del HTLV-I comparten los mismos pesos moleculares, las bandas de WB del HTLV-I aparentarán ser positivas al VIH.
Un problema más directo asociado con el uso del "aislamiento del VIH" como estándar oro es el hecho de que, independientemente de los distintos fenómenos aceptados por los investigadores del sida como representantes del "aislamiento del VIH", y a pesar de que no se han escatimado esfuerzos, no es posible "aislar el VIH" a partir de todos los pacientes positivos de anticuerpos. El ratio de éxito varía entre el 17% y el 80% (92,93,123).
Al contrario, cuando se realiza el mismo esfuerzo, el VIH se puede aislar a partir de algunos pacientes seronegativos sin sida, e incluso de individuos seronegativos normales sin riesgo de infección por VIH (124,125). Con un método más reciente utilizado para el "aislamiento del VIH", la detección de la p24 en cultivos con sangre completamente sin fraccionar (126,127), se informó de resultados positivos en 49 de 60 (82%) de los "individuos presumiblemente no infectados pero serológicamente indeterminados" y en 5 de 5 "donantes de sangre seronegativos" (128).
Ya en 1988, investigadores del CDC en los Estados Unidos se dieron cuenta que no existe correlación entre el "aislamiento del VIH" y un test de anticuerpos positivo (que ellos llaman infección documentada), y más importante, entre "aislamiento del VIH" in vitro y su presencia in vivo: "la correlación entre estos dos métodos es limitada; son inconsistentes, en que el virus no puede detectarse en cada persona con una infección documentada. Además, la técnica de cultivo determina la capacidad de las células infectadas de producir virus in vitro, pero no indica necesariamente el estado de expresión de virus in vivo". (129).
Investigaciones genómicas
En las décadas siguientes a los experimentos de Rous, Rous así como otros investigadores llevaron a cabo investigaciones similares con algunas especies animales. Sin embargo, aunque la neoplasia podría inducirse mediante la inyección de filtrados de tejidos de tumor (retrovirus infecciosos, retrovirus exógenos), no hubo datos epidemiológicos para sugerir un origen infeccioso del cáncer.
En 1939 Andrews "especuló en la posible activación de partículas infecciosas virales latentes en teijdos cancerosos", y en 1948 Darlington propuso "que tales virus [virus endógenos] podrían surgir de elementos genéticos celulares que llamó provirus" (80).
En los años 50 y 60, los siguientes datos experimentales se consideraron como prueba de la hipótesis proviral: (a) animales sanos en los cuales ningún virus completo podía detectarse, tenían antígenos virales similares a los de los virus exógenos; (b) genomas de ADN o genomas parciales de los retrovirus infecciosos, se encontró que estaban integrados en los genomas de celulas normales que no producen virus; (c) "La prueba final vino con el aislamiento de virus infecciosos a partir de células no infectadas". Se encontró que cuando eran cultivadas, las células sanas que no producen virus sí producían virus espontáneamente (80). Su aparición y producción podrían incrementarse un millón de veces mediante: (i) estimulación mitogénica (130); (ii) técnicas de cocultivación (131); (iii) cultivación de células con sobrenadante procedente de cultivos de producción no viral (132). (Nota: la estimulación mitogénica es un requisito indispensable para el aislamiento del VIH, y de hecho, en la mayoría de los casos, se emplean todos los anteriores).
En la actualidad se acepta generalmente que "una de las características más llamativas que distingue a los retrovirus de todos los demás virus animales es la presencia, en los cromosomas de células no infectadas normales, de genomas estrechamente relacionados con, o idénticos a, los de los virus infecciosos". (80)
Dependiendo de las condiciones, el genoma de provirus permanece sin expresar, o puede expresarse todo o parte del mismo. Cuando se expresa todo, esto puede llevar o no al ensamblaje de partículas virales (retrovirus endógeno) (80). Es decir, el hallazgo de un genoma viral (ADN), o incluso de ARN, antígenos y anticuerpos a los mismos, no es prueba de la presencia de partículas infecciosas.
Aunque la mayoría de los hallazgos anteriores son de experimentos con animales, en la actualidad existen datos referentes a que "El genoma humano lleva secuencias de ADN relacionadas con genomas retrovirales endógenos que están divididas en familias según la homología de secuencia. Algunas están presentes en solamente unas copias, mientras otras están presentes desde cientos a miles de copias" (133).
Los datos en animales también muestran que pueden surgir nuevos retrovirus mediante mezcla fenotípica, y recombinación y borrado genético.
Cuando una célula contiene dos provirus, se puede encontrar descendencia que posee el genoma de uno, y las proteínas estructurales de cualquiera o ambos de los virus presentes. Al contrario, el ARN puede ser viral, pero al menos algunas de las proteínas pueden ser celulares.
En otros casos, las partículas no tienen un genoma en absoluto, o faltan uno o más genes (virus genéticamente defectuosos). La mezcla genética puede ser entre genomas virales o entre genes virales y celulares (80,134).
Según distinguidos retrovirólogos como Weiss y Temin, pueden surgir nuevos genomas retrovirales mediante reensamblaje de ADN celular, algo causado por muchos factores, incluyendo procesos patogénicos, una visión que propone los retrovirus como un efecto, y no la causa, de la enfermedad (135,136).
El tiempo y aparición del genoma viral "puede ser millones de años en células de línea germinal, y días en las células somáticas" (136).
Además de lo anterior, el ciclo de replicación retroviral "conlleva tres pasos distintos: transcripción inversa, polimerización de ADN y la síntesis de ARN a partir de un patrón de ADN (transcripción). Cualquier error cometido por la enzima de polimerasa durante el primer y el tercer paso no está sujeto a lectura de pruebas, siendo el resultado variabilidad de secuencia pronunciada" (137).
De este modo, tan pronto como en 1973, se concluyó que los anteriores fenómenos "constituirán un obstáculo para cualquier análisis genético de virus de tumor de ARN" (138) (virus de tumor de ARN = retrovirus). Hasta el día de hoy, los datos sobre el genoma del VIH no han modificado la predicción anterior, y muestran que podrían existir muchos problemas con el uso de los estudios genómicos en esfuerzos para probar la infección de los pacientes de sida con un retrovirus exógeno singular, el VIH.
Algunos de estos problemas se pueden resumir como sigue:
(I) No hay dos genomas del VIH que sean el mismo. (a) No se han aislado dos VIH idénticos incluso de la misma persona. En un caso, donde se hicieron dos aislamientos secuenciales separados en 16 meses, ninguno de los provirus en el primer aislamiento se encontró en el segúndo (139), llevando a un investigador del VIH a concluir que "Los datos implican que no hay tal aislamiento [virus del sida]" (140); (b) de una misma persona en un momento determinado se puede aislar más de un VIH (141,142); (c) muchos, si no todos, de los provirus detectados in vitro e in vivo son defectuosos (143); (d) en un mismo paciente, los datos genómicos en monocitos difieren de aquellos en linfocitos T (144); (e) los datos genéticos obtenidos in vitro no correlan con los datos obtenidos in vivo: "cultivar es perturbar" (145); (f) El tipo de virus aislado se determina por los tipos de células utilizados para el aislamiento de VIH (142,146).
(II) No hay correlación entre el "aislamiento" del VIH y la detección del genoma del VIH. Los cultivos positivos en "infección de virus" pueden ser "negativos en reacción en cadena de la polimerasa" (147). (III) Las secuencias del VIH no se encuentran en todos los pacientes de sida. Gallo y sus colegas, al resumir el primer estudio de hibridación con tejido fresco concluyeron: "Hemos sido capaces anteriormente de aislar el HTLV-III a partir de sangre periférica y tejido de ganglio linfático procedente de la mayoría de pacientes con sida o ARC" [aproximadamente el 50% de los pacientes a los que se refiere Gallo]. "Sin embargo, como se muestra aquí, el ADN del HTLV-III normalmente no se detecta mediante hibridación Southern Blotting de estos mismos tejidos y, cuando sí se detecta, las bandas son con frecuencia débiles ... el aumento del ganglio linfático comunmente encontrado en pacientes de ARC y sida puede no deberse directamente a la proliferación de células infectadas con el HTLV-III ... la ausencia de secuencias HTLV-III detectables en tejidos de sarcoma de Kaposi de pacientes de sida sugiere que este tumor no está directamente causado por la infección de cada célula de tumor por el HTLV-III ... la observación de que las secuencias de HTLV-III se encuentran raramente, o para nada, en células mononucleares de sangre periférica, médula osea, y bazo proporcionan los primeros datos diretos de que estos tejidos no están fuerte o ampliamente infectados con el HTLV-III ni en el sida ni en el ARC" (148). Estos estudios fueron confirmados por muchos otros investigadores.
Para mejorar la detección se introdujo el método de la reacción en cadena de la polimerasa (PCR). Sin embargo, "una característica llamativa de los resultados obtenidos hasta ahora" con este método, al igual que con la técnica de hibridación estándar, "es la escasez o ausencia aparente de ADN viral en una proporción de pacientes" (149) y, cuando se encuentra ARN o ADN viral, la "señal" es muy pequeña.
Por ejemplo, se piensa que el VIH se transmite principalmente por relaciones sexuales, aunque, con la técnica PCR, el "genoma del VIH" se puede detectar en una minoría de muestras de semen (1 de 25) (147). Debe señalarse que un positivo PCR, incluso si se encuentra en todos los pacientes, como se afirma en algunas publicaciones (149), no puede considerarse que signifique la presencia del genoma completo del VIH. Con la PCR "solamente pueden amplificarse pequeñas regiones, un gen lo más" (143), es decir, no se detecta el genoma viral completo y, puesto que la mayoría de los "aislamientos" de VIH hasta la fecha son defectuosos, la detección de parte de un gen o de un gen completo, o incluso de algunos genes, no se puede considerar sinónimo del genoma de VIH completo.
Además, la PCR no está estandarizada y, hasta la fecha, ha habido un único estudio en donde se examinó la reproducibilidad, sensitividad y especificidad de la PCR. En este estudio, el estándar oro que se utilizó fue el estado serológico. La especificidad se determinó midiendo el porcentaje de resultados PCR negativos en individuos de bajo riesgo sanos (donantes de sangre) seronegativos (ELISA).
La PCR resultó no ser reproducible, y "se observaron resultados falsos positivos y falsos negativos en todos los laboratorios (la concordancia con la serología varió del 40% al 100%). Además, el número de resultados PCR positivos no se diferenció significativamente entre seronegativos de alto y bajo riesgo" (150).
(IV) Los resultados de hibridación positiva podrían no ser específicos al VIH. En 1984, cuando Gallo y sus colegas condujeron los primeros estudios de hibridación, encontraron que cuando los resultados eran positivos, las bandas hibridación eran "débiles", "señal baja".
La "señal baja" se interpretó como prueba de que los individuos infectados con el VIH contienen provirus en números pequeños de células mononucleares de sangre periférica, y en números de copia bajos. Sin embargo, según Gallo y sus colegas "teoréticamente esta intensidad de señal baja podría también explicarse por la presencia de un virus lejánamente homólogo al HTLV-III en estas células" (148). Los datos que han salido a la luz desde entonces sugieren que esta posibilidad teórica podría ser un hecho: (a) Aunque ya no se acepta que el VIH sea transmitido por insectos, en 1986 investigadores del Instituto Pasteur encontraron secuencias de ADN de VIH en moscas tse-tse, escarabajos negros y en leones de Zaire y la República Centroafricana (151). (b) En 1984 el grupo de Gallo reportó que el genoma del VIH se hibrida con los genes estructurales (gag, pol y env) de tanto el HTLV-I como el HTLV-II" (152). Los datos disponibles actualmente muestran que el ADN normal humano contiene secuencias genómicas retrovirales relacionadas con el HTLV-I y HTLV-II (153,154). (c) En 1985 Weiss y sus colegas reportaron el aislamiento, a partir de cultivos de células T estimulados mitogénicamente procedentes de dos pacientes con hipogammaglobulinemia variable común, de un retrovirus que "estaba claramente relacionado con el HTLV-III/LAV"; los datos incluyeron un WB positivo con sueros de sida y la hibridación con sondas de VIH (155). (d) el ADN extraído de glándulas tiroides de pacientes con la enfermedad de Grave hibridan con "la region de codificación gag p24 completa" del VIH (156). (e) Horowitz et al "describen el primer informe de la presencia de secuencias de nucleótidos relacionados con el VIH-1 en ADN's de humanos, de chinpancés y de monos Rhesus procedentes de individuos no infectados normales". Ellos "demostraron la presencia de una familia compleja de secuencias relacionadas con el VIH-1" en las anteriores especies, y concluyeron que "un análisis adicional de los miembros de esta familia ayudará a determinar si tales secuencias endógenas contribuyeron a la evolución del VIH-1 mediante eventos de recombinación, o si estos elementos, bien directamente o a traves de productos de proteínas, influencian la patogénesis del VIH" (157).
El que las señales de hibridación positivas pueden deberse a que tales eventos sean causados por los agentes oxidantes (mutágenos y mitógenos) a los que los grupos del riesgo del sida y los cultivos están expuestos, se sugiere por lo siguiente: Un PCR positivo revierte a negativo cuando la exposición a factores de riesgo cesa (158,159), y los monocitos de pacientes VIH positivos en que no se puede detectar ADN del VIH, ni siquiera mediante la PCR, llegan a ser positivos de ARN de VIH tras la cocultivación con células T activadas ConA normales" (160). Ya en 1989 investigadores del Instituto Pasteur concluyeron que "la tarea de definir la infección por VIH en términos moleculares será difícil" (145). Confirmaron su conclusión en un estudio reciente donde "describieron la enorme heterogeneidad encontrada in vivo dentro de las poblaciones del VIH-1", y la posibilidad "de que un portador del VIH pueda fácilmente albergar más de 1.010 provirus, la mayoría de los cuales será geneticamente singular". Concluyeron: "Es por tanto posible que el tamaño puro de las variantes dentro de un individuo infectado permita al VIH explorar posibilidades genéticas totalmente nuevas". La aparición de estructuras retrovirales "genéticas radicalmente distintas" podría ser el resultado de "reensamblaje, duplicación, borrado o hipermutación. La transducción del ADN de la célula huésped representa posiblemente el rasgo genético más sorprendente de los retrovirus" (161).
Conclusión
Es axiomático que el uso de tests de anticuerpos debe verificarse frente a un estándar oro. Los datos disponibles actualmente no proporcionan tal estándar oro para los tests de anticuerpos del VIH. La conclusión ineludible del anterior análisis es que el uso de los tests de anticuerpos del VIH como herramientas predictivas, diagnósticas y epidemiológicas de la infección de VIH necesita ser cuidadosamente reevaluado.*
Agradecimientos
Nos gustaría expresar nuestro agradecimiento a todos nuestros colegas, y especialmente a Udo SchEklenk, Barry Page, Bruce Hedland-Thomas, David Causer, Richard Fox, John Peacock, David Prentice, Ronald Hirsch, Patricia Shalala, Keith Jones, Alun Dufty, June Rider Jones, Coronary Barrow, Dorothy Davis, Julian Smith, Mark Strahan, Vincent Turner, Wallace Turner y Graham Drabble por su continuo apoyo y ayuda.
Dedicación
Este trabajo está dedicado a la memoria de Methodios Papadopulos y Margaret Joan Turner.
Autores:
Eleni Papadopulos-Eleopulos, Physicist
Department of Medical Physics
Royal Perth Hospital
Valendar F. Turner, Staff Specialist
Department of Emergency Medicine
Royal Perth Hospital
John M. Papadimitriou Professor of Pathology
Department of Pathology
University of Western Australia
Correspondencia a:
Eleni Papadopulos-Eleopulos
Department of Medical Physics
Royal Perth Hospital
Box X2213 GPO Perth
Referencias
2. Hausmann, E.H.S., Gelderblom, H.R., Clapham, P.R. et al. 1987. Detection of HIV envelope specific antibodies by immunoelectron microscopy and correlation with antibody titer and virus neutralizing activity. J. Virol. Meth. 16:125-137.
3. Pinter, A., Honnen, W.J., Tilley, S.A. et al. 1989. Oligomeric Structure of gp41,the Transmembrane Protein of Human Immunodeficiency Virus Type 1. J. Virol. 63:2674-2679.
4. BarrG-Sinoussi, F., Chermann, J.C., Rey, F. et al. 1983. Isolation of a T-Lymphotrophic Retrovirus from a patient at Risk for Acquired Immune Deficiency Syndrome (AIDS). Science 220:868-871.
5. SchEpbach, J., Popovic, M., Gilden, R.V. et al. 1984. Serological analysis of a Subgroup of Human T-Lymphotrophic Retroviruses (HTLV-III) Associated with AIDS. Science 224:503-505.
6. Damsky, C.H., Sheffield, J.B., Tuszynski, G.P. et al. 1977. Is there a role for Actin in Virus Budding? J. Cell. Biol. 75:593-605.
7. Stanislawsky, L., Mongiat, F., Neto, V.M. et al 1984. Presence of Actin in Oncornaviruses. Biochem. Biophys. Res. Com. 118:580-586.
8. Papadopulos-Eleopulos, E., Turner, V.F. and Papadimitriou, J.M. 1992. Oxidative stress, HIV and AIDS. Res. Immunol. 143:145-148.
9. Hinshaw, D.B., Burger, J.M., Beals, T.F. et al. 1991. Actin polymerization in cellular oxidant injury. Arch. Biochem. Biophys. 228:311-316.
10. Bach, M.A., Lewis, D.E., McClure, J.E. et al. 1986. Monoclonal Anti-actin Antibody Recognizes a Surface Molecule on Normal and Transformed Human B Lymphocytes:Expression Varies with Phase of Cell Cycle. Cell. Immunol. 98:364-374.
11. Stricker, R.B., Abrams, D.I., Corash, L. et al. 1985. Target Platelet Antigen in Homosexual Men with Immune Thrombocytopenia. NEJM 313:1375-1380.
12. Henderson, L.E., Sowder, R., Copeland, T.D. et al. 1987. Direct Identification of Class II Histocompatibility DR Proteins in Preparations of Human T-Cell Lymphotropic Virus Type III. J. Virol. 61:629-632.
13. Wong-Staal, F. and Gallo, R.C. 1985. Human T-lymphotropic retroviruses. Nature 317:395-403.
14. Genesca, J., Jett, B.W., Epstein, J.S. et al. 1989. What do Western Blot indeterminate patterns for Human Immunodeficiency Virus mean in EIA-negative blood donors? Lancet II:1023-1025.
15. Ranki, A., Johansson, E. and Krohn, K. 1988. Interpretation of Antibodies Reacting Solely with Human Retroviral Core Proteins. NEJM 318:448-449.
16. Delord, B., Ottmann, M., Schrive, M.H. et al. 1991. HIV-1 expression in 25 infected patients:A comparison of RNA PCR, p24 EIA in Plasma and in situ Hybridization in mononuclear cells, p113. In: Vol. I, Abstracts VII International Conference on AIDS,Florence.
17. Todak, G., Klein, E., Lange, M. et al. 1991. A clinical appraisal of the p24 Antigen test, p326. In:Vol. I, Abstracts VII International Conference on AIDS,Florence.
18. Courouce, A., Muller, J. and Richard, B. 1986. False-Positive Western Blot Reactions to Human Immunodeficiency Virus in Blood Donors. Lancet II:921-922.
19. Stricker, R.B., McHugh, T.M., Moody, D.J. et al. 1987. An AIDS-related Cytotoxic autoantibody reacts with a specific antigen on stimulated CD4+ T cells. Nature 327:710-713.
20. Chassagne, J., Verelle, P., Fonck, Y. et al. 1986. Detection of the Lymphadenopathy-Associated Virus p18 in cells of patients with Lymphoid Diseases using a Monoclonal Antibody. Ann. Inst. Pasteur/Immunol. 137D:403-408.
21. Parravicini, C.L., Klatzmann, D., Jaffray, P. et al. 1988. Monoclonal Antibodies to the human immunodeficiency virus p18 protein cross-react with normal human tissues. AIDS 2:171-177.
22. Matsiota, P., Chamaret, S., Montagnier, L. et al. 1987. Detection of Natural Autoantibodies in the serum of Anti-HIV Positive-Individuals. Ann. Inst. Pasteur/Immunol. 138:223-233.
23. Popovic, M., Sarngadharan, M.G., Read, E. et al 1984. Detection, Isolation,and Continuous Production of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and Pre-AIDS. Science 224:497-500.
24. Wilber, J.C. 1991. New Developments in Diagnosing Infections, p.1-15. In:AIDS Clinical Review, P. Volbering, M.A. Jacobson (Eds.). Marcel Dekker Inc. New York.
25. Lundberg, G.D. 1988. Serological Diagnosis of Human Immunodeficiency Virus Infection by Western Blot Testing. JAMA 260:674-679.
26. Zolla-Pazner, S., Gorny, M.K. and Honnen, W.J. 1989. Reinterpretation of Human Immunodeficiency Virus Western Blot Patterns. NEJM 320:1280-1281.
27. Burke, D.S. 1989. Laboratory Diagnosis of Human Immunodeficiency Virus Infection. Clin. Lab. Med. 9:369-392.
28. Maskill, W.J. and Gust, I.D. 1992. HIV-1 Testing in Australia. Australian Prescriber 15:11-13.
29. DeCock, K.M., Selik, R.M., Soro, B. et al. 1991. AIDS surveillance in Africa:a reappraisal of case definitions. BMJ 303:1185-1189.
30. Voevodin, A. 1992. HIV screening in Russia. Lancet 339:1548.
31. Working Group on AIDS Case Definition 1990. Epidemiol. Bull. 4:9-11.
32. Edwards, V.M., Mosley, J.W. and the Transfusion Safety Study Group. 1991. Reproducibility in Quality Control of Protein (Western) Immunoblot Assay for Antibodies to Human Immunodeficiency Virus. Am. J. Clin. Pathol. 91:75-78.
33. CDC. 1989. Interpretation and Use of the Western Blot Assay for Serodiagnosis of Human Immunodeficiency Virus Type 1 Infections. MMWR 38 No. S-7:1-7.
34. Weiss, S.H., Goedert, J.J., Sarngadharan, M.G. et al. 1985. Screening Test for HTLV-III (AIDS Agent) Antibodies. JAMA 253:221-225.
35. Weiss, R. and Thier, S.O. 1988. HIV testing is the answer-what's the question? NEJM 319:1010-1012.
36.Burke, D.S., Brundage, J.F., Redfield, R.R. et al. 1988. Measurement of the False Positive Rate in a Screening Program for Human Immunodeficiency Virus Infections. NEJM 319:961-964.
37. Scarlatti, G.S., Lombardi, V., Plebani, A. et al. 1991. Polymerase chain reaction, virus isolation and antigen assay in HIV-1-antibody-positive mothers and their children. AIDS 5:1173-1178.
38. Griner, P.F., Mayewski, R.J., Mushlin, A.I. et al. 1981. Selection and Interpretation of Diagnostic Tests and Procedures. Ann. Int. Med. 94 (Part 2):559-563.
39. CDC. 1987. Revision of the CDC Surveillance Case Definition for Acquired Immunodeficiency Syndrome. JAMA 258:1143-1145.
40. Conley, C.L. and Savarese, D. 1989. Biologic False-Positive Serologic Tests for Syphilis and Other Serologic Abnormalities in Autoimmune Hemolytic Anemia and Thromobocytopenic Purpura. Medicine 68:67-84.
41. Boue, F., Dreyfus, M., Bridley, F. et al. 1990. Lupus anticoagulant and HIV infection:a prospective study. AIDS 4:467-471.
42. Jaffe, J.H., Moore, J.D., Cone, E.J. et al. 1986. HTLV-III Seropositivity in 1971-1972 Parenteral Drug Abusers-A case of false Positives or Evidence of Viral Exposure? NEJM 314:1387-1388.
43. Biggar, R.J., Gigase, P.L., Melbye, M. et al. 1985. ELISA HTLV Retrovirus Antibody Reactivity Associated with Malaria and Immune Complexes in Healthy Africans. Lancet II:520-523.
44. Ternynck, T. and Avrameas, S. 1986. Murine Natural Monoclonal Autoantibodies: A Study of their Polyspecificities and their Affinities. Immunol. Rev. 94:99-112.
45. Pateraki, E., Kaklamani, E., Portocalas, K.R. et al. 1986. Autoantibodies in systemic lupus erythematosus and normal subjects. Clin. Rheumatol. 5:338-345.
46. Morton, D.L. and Malmgren, R.A. 1968. Human Osteosarcomas:Immunologic Evidence suggesting an associated agent. Science 162:1279-1281.
47. Hirshaut, Y., Pei, D.T., Marcove, R.C. et al. 1974. Seroepidemiology of Human Sarcoma Antigen (S1). NEJM 291:1103-1107.
48. Kurth, R., Teich, N.M., Weiss, R. et al. 1977. Natural human antibodies reactive with primate type-C viral antigens. Proc. Natl. Acad. Sci. 74:1237-1241.
49. Snyder, H.W. and Fleissner, E. 1980. Specificity of human antibodies to oncovirus glycoproteins:Recognition of antigen by natural antibodies directed against carbohydrate structures. Proc. Natl. Acad. Sci. 77:1622-1626.
50. Barbacid, M., Bolognesi, D. and Aaronson, S.A. 1980..Humans have antibodies capable of recognizing oncoviral glycoproteins:Demonstration that these antibodies are formed in response to cellular modification of glycoproteins rather than as consequence of exposure to virus. Proc. Natl. Acad. Sci. 77:1617-1621.
51. Essex, M., McLane, M.F., Lee, T.H. et al. 1983. Antibodies to Cell Membrane Antigens Associated with Human T-Cell Leukemia Virus in Patients with AIDS. Science 220:859-862.
52. Novick, D.M., Des Jarlais, D.C., Kreek, M.J. et al. 1988. Specificity of Antibody Tests for Human Immunodeficiency Virus in Alcohol and Parenteral Drug Abusers with Chronic Liver Disease. Alcoholism Clin Exp Res 12:687-690.
53. European Collaborative Study. 1991. Children born to women with HIV-1 infection:natural history and risk of transmission. Lancet 337:253-260.
54. Beneviste, R.E., Ochs, H.D., Fischer, S.H. et al. 1986. Screening for antibodies to LAV/HTLV-III in recipients of immunoglobulin preparations. Lancet I:1090-1092.
55. Burinsky, K.I., Chaplinskas, S.A., Syrtsev, V.A. et al. 1988. Reactivity to gag- and env-related sequences in immunoblot assay is not necessarily indicative of HIV infection. AIDS 2:405-406.
56. Calabrese, L.H. 1988. Autoimmune Manifestations of Human Immunodeficiency Virus (HIV) Infection. Clin. Lab. Med. 8:269-279.
57. Bonara, P., Maggioni, L. and Colombo, G. 1991. Anti-Lymphocyte Antibodies and Progression of Disease in HIV Infected Patients,p.149. In:Vol. II, VII International Conference on AIDS,Florence.
58. Tumietto, F., Costigliola, P., Ricchi, E. et al. 1991. Anti-Lymphocyte Auto-antibodies:Evaluation and correlation with different stages of HIV Infection,p149. In:Vol. II, VII International Conference on AIDS,Florence.
59. Morozov, V.A., Ilyinskii, P.O., Uckert, W.A. et al. 1989. Antibodies to structural and nonstructural gag-coded proteins of type-D retroviruses in human with lymphadenopathy and AIDS. Int. J. Tiss. Reac. XI:1-5.
60. Lucey, D.R., Hendrix, C.W., Andrzejewski, C. et al. 1992. Comparison by Race of Total Serum IgG,IgA,and IgM with CD4+ T-Cell Counts in North American Persons Infected with the Human Immunodeficiency Virus Type 1. J. Acquir. Immun. Defic. Syndr. 5:325-332.
61. Papadopulos-Eleopulos, E. 1988. Reappraisal of AIDS:Is the oxidation induced by the risk factors the primary cause? Med. Hypotheses 25:151-162
62. Papadopulos-Eleopulos, E., Turner, V.F. and Papadimitriou, J. 1992. Kaposi's sarcoma and HIV. Med. Hypotheses 39:22-29.
63. Igel, H.J., Turner, H.C., Kotin, P. et al. 1969. Mouse Leukaemia Virus Activation by Chemical Carcinogens. Science 166:624-1626.
64.Sterk, C. 1988. Cocaine and HIV Seropositivity. Lancet I:1052-1053.
65. Kion, T.A. and Hoffmann, G.W. 1991. Anti-HIV and Anti-Anti-MHC Antibodies in Alloimmune and Autoimmune Mice. Science 253:1138-1140.
66. Conley, L.J. and Holmerg, S.D. 1992. Transmission of AIDS from blood screened negative for antibody to the human immunodeficiency virus. NEJM 326:1499.
67. Burgher, H., Weiser, B., Robinson, W.S. et al. 1985. Transient Antibody to Lymphadenopathy-Associated/Human T-Lymphotrophic Virus Type III and T-Lymphocyte Abnormalities in the Wife of a Man who Developed the Acquired Immunodeficiency Syndrome. Ann. Int. Med. 103:545-547.
68. Esteva, M.H., Blasini, A.M., Ogly, D. and Rodriguez, M.A. 1992. False positive results from antibody to HIV in two men with systemic lupus erythrematosus. Ann. Rhem. Dis. 51:1071-1073.
69. Dummer, J.S., Erb, S., Breinig, M.K. et al. 1989. Infection with Human Immunodeficiency Virus in the Pittsburgh Transplant Population. Transplantation 47:134-139.
70. Pitchenik, A.E., Burr, J., Suarez, M. et al. 1987. Human T-Cell Lymphotrophic Virus-III (HTLV-III) Seropositivity and Related Disease Among 71 Consecutive Patients in Whom Tuberculosis was Diagnosed. Am. Rev. Respir. Dis. 135:875-879.
71. Nzilambi, N., Mann, J.M., Francis, H. et al. 1986. Seroprevalence among tuberculosis patients in Zaire. In:Abstracts II International AIDS Conference, Paris, No.105:S17b.
72. St.Louis, M.E., Rauch, K.J., Peterson, L.R. et al. 1990. Seroprevalence rates of Human Immunodeficiency Virus Infection at Sentinel Hospitals in the United States. NEJM 323:213-218.
73. Chamberland, M., Conley, L. and Dondero, T. 1988. Epidemiology of heterosexually acquired AIDS-United States, p.264. In:Abstacts IV International AIDS Conference,Stockholm. 74. New York City AIDS Surveillance Report, January-March 1992.
75. Rodriquez, L., Dewhurst, S., Sinangil, F. et al. 1985. Antibodies to HTLV-III/LAV among Aboriginal Amazonian Indians in Venezuela. Lancet II:1098-1100.
76. Volsky, D.J., Wu, Y.T., Stevenson, M. et al 1986. Antibodies to HTLV-III/LAV in Venezuelan patients with acute malarial syndromes. NEJM 314:647.
77. Rous, P. 1911. A Sarcoma of the Fowl transmissible by an agent separable from the Tumor Cells. J. Exp. Med. 13:397-411.
78. Toplin, I. 1973. Tumor Virus Purification using Zonal Rotors. Spectra No. 4:225-235.
79. Temin, H.M. and Baltimore, D. 1972. RNA-Directed DNA Synthesis and RNA Tumor Viruses. Adv. Vir. Res. 17:129-186.
80. RNA Tumor Viruses. 1982. R. Weiss, N. Teich, H. Varmus, J. Coffin (Eds.).Cold Spring Harbor Laboratory. Cold Spring Harbor, New York.
81. Bader, J.P. 1975. Reproduction of RNA Tumor Viruses, p.253-331. In:Comprehensive Virology Vol.4. H. Fraenkel-Conrat, R.R. Wagner (Eds.). Plenum Press, New York.
82. Sinoussi, F., Mendiola, L., Chermann, J.C. et al. 1973. Purification and partial differentiation of the particles of murine sarcoma virus (M. MSV) according to their sedimentation rates in sucrose density gradients. Spectra No. 4:237-243.
83. Amiesen, J.C. and Capron, A. 1991. Cell dysfunction and depletion in AIDS: the programmed cell death hypothesis. Immunol. Today 12:102-105.
84. Wyllie, A.H., Kerr, J.F.R. and Currie, A.R. 1980. Cell Death:The Significance of Apoptosis. Int. Rev. Cytol. 68:252-306.
85. Hoxie, J.A., Fitzharris, T.P., Youngbar, P.R. et al. 1987. Nonrandom Association of Cellular Antigens with HTLV-III Virions. Hum. Immunol. 18:39-52.
86. McKeating, J.A. and Moore, J.P. 1991. HIV infectivity. Nature 349:660.
87. Masquelier, B., Combeau, T. and Poveda, J. 1991. In Vitro Assays Show a Dissociation of Reverse Transcriptase Activity and Core Antigen (p24) Production in Two HIV-1 Isolates from a Patient Receiving Long-Term Treatment with Zidovudine (ZDV). J. Acquir. Immun. Defic. Syndr. 4:499-505.
88. Grunow, R., Valentin, A., Fenyo, E.M. et al. 1991. Release of HIV-1 core protein from infected cells independent of infectious virus particles, p.157. In:Vol. I, Abstracts VII International Conference on AIDS,Florence.
89. Tedder, R.S., Semple, M.G., Tenant-Flowers, M. et al. 1992. HIV, AIDS, and zidovudine. Lancet 339:805-806.
90. Lemaetre, M., GuGtard, D., HGnin, Y. et al. 1990. Protective Activity of Tetracycline Analogs against the Cytopathic effect of the Human Immunodeficiency Viruses in CEM Cells. Res. Virol. 141:5-16.
91. Boulerice, F., Bour, S., Geleziunas, R. et al. 1990. High Frequency of Isolation of Defective Human Immunodeficiency Virus Type 1 and Heterogeneity of Viral Gene Expression in Clones of Infected U-937 Cells. J. Virol. 64:1745-1755.
92. Ehrnst, A., Sonnerborg, A., Bergdahl, S. and Strammegard, O. 1988. Efficient Isolation of HIV From Plasma During Different Stages of HIV Infection. J. Med. Virol. 26:23-32.
93. Learmont, J., Tindall, B., Evans, L. et al. 1992. Long-term symptomless HIV-1 infection in recipients of blood products from a single donor. Lancet 340:863-867.
94. Popovic, M., Sarngadharan, M.G., Read, E. et al. 1984. Detection, Isolation, and Continuous Production of Cytopathic Retroviruses (HTLV-III) from Patients with AIDS and Pre-AIDS. Science 224:497-500.
95. Gallo, R.C., Sarin, P.S. and Wu, A.M. 1973. On the nature of the Nucleic Acids and RNA Dependent DNA Polymerase from RNA Tumor Viruses and Human Cells, p.13-34. In:Possible Episomes in Eukaryotes. L.G. Silvestri (Ed.). North-Holland Publishing Company, Amsterdam.
96. Whitkin, S.S., Higgins, P.J. and Bendich, A. 1978. Inhibition of reverse transcriptase and human sperm DNA polymerase by anti-sperm antibodies. Clin. Exp. Immunol. 33:244-251.
97. Tomley, F.M., Armstrong, S.J., Mahy, B.W.J. and Owen, L.N. 1983. Reverse transcriptase activity and particles of retroviral density in cultured canine lymphosarcoma supernatants. Br. J. Cancer 47:277-284.
98. Weissbach, A., Baltimore, D., Bollum, F. et al. 1975. Nomenclature of Eukaryotic DNA Polymerases. Science 190:401-402.
99. Gallo, R.C., Wong-Staal, F., Reitz, M. et al. 1976. Some Evidence For Infectious Type-C Virus in Humans, p.385-407. In:Animal Virology. D. Baltimore, A. S. Huang, C.F. Fox (Eds.). Academic Press Inc., New York.
100. Panem, S., Prochownik, E.V., Reale, F.R. et al. 1975. Isolation of Type C Virions from a Normal Human Fibroblast Strain. Science 189:297-299.
101. Panem, S., Prochownik, E.V., Knish, W.M. and Kirsten, W.H. 1977. Cell Generation and Type-C Virus Expression in the Human Embryonic Cell Strain HEL-12. J. Gen. Virol. 35:487-495.
102. Panem, S. 1979. C Type Virus Expression in the Placenta. Curr. Top. Pathol. 66:175-189.
103. Sarngadharan, M.G., Robert-Guroff, M. and Gallo, R.C. 1978. DNA Polymerases of Normal and Neoplastic Mammalian Cells. Biochim. Biophys. Acta. 516:419-487.
104. Klatzmann, D., BarrG-Sinoussi, F., Nugeyre, M.T. et al. 1984. Selective Tropism of Lymphadenopathy Associated Virus (LAV) for Helper-Inducer T Lymphocytes. Science 225:59-63.
105. Montagnier, L. 1985. Lymphadenopathy-Associated Virus:From Molecular Biology to Pathogenicity. Ann. Int. Med. 103:689-693.
106. Gendelman, H.E., Orenstein, J.M., Martin, M.A. et al 1988. Efficient Isolation and Propagation of Human Immunodeficiency Virus on Recombinant Colony-Stimulating Factor 1-Treated Monocytes. J. Exp. Med. 167:1428-1441.
107. Gelderblom, H.R., Hausmann, E.H.S., tzel, M. et al. 1987. Fine Structure of Human Immunodeficiency Virus (HIV) and Immunolocalization of Structural Proteins. Virol. 156:171-176.
108. Gelderblom, H., Reupke, H., Winkel, T. et al. 1987. MHC-Antigens:Constituents of the Envelopes of Human and Simian Immunodeficiency Viruses. Z. Naturforsch. 42C:1328-1334.
109. Gelderblom, H.R., Reupke, H. and Pauli, G. 1985. Loss of envelope antigens of HTLV-III/LAV, a factor in AIDS pathogenesis? Lancet II:1016-1017.
110. Neidrig, M., Rabanus, J.P., L'Age Stehr, J. et al. 1988. Monoclonal Antibodies Directed against Human Immunodeficiency Virus (HIV) gag Proteins with Specificity for Conserved Epitopes in HIV-1, HIV-2 and Simian Immunodeficiency Virus. J. Gen. Virol. 69:2109-2114.
111. Gelderblom, H.R., tzel, M., Hausmann, E.H.S. et al. 1988. Fine Structure of Human Immunodeficiency Virus (HIV),Immunolocalization of Structural Proteins and Virus-Cell Relation. Micron Microsc. 19:41-60.
112. Meerloo, T., Parmentier, H.K., Osterhaus, A. et al. 1992. Modulation of cell surface molecules during HIV-1 infection of H9 cells. An immunoelectron microscopic study. AIDS 6:1105-1116.
113. Lecatsas, G. and Taylor, M.B. 1986. Pleomorphism in HTLV-III,the AIDS virus. S. Afr. Med. J. 69:793-794.
114. Hockley, D.J., Wood, R.D., Jacobs, J.P. et al. 1988. Electron Microscopy of Human Immunodeficiency Virus. J. Gen.Virol. 69:2455-2469.
115. Dourmashkin, R.R., O'Toole, C.M., Bucher, D. and Oxford, J.S. 1991.The presence of budding virus-like particles in human lymphoid cells used for HIV cultivation. p.122. In:Vol. I, Abstracts VII International Conference on AIDS,Florence.
116. Brennan, J.K., Lichtman, M.A., Chamberlain, J.K. and Leblond, P. 1976. Isolation of Variant Lymphoma Cells with Reduced Growth Requirements for Extracellular Calcium and Magnesium and Enhanced Oncogenicity. Blood 47:447-459.
117. Garry, R.F., Fermin, C.D., Hart, D.J. et al. 1990. Detection of a Human Intracisternal A-Type Retroviral Particle Antigenically Related to HIV. Science 250:1127-1129.
118. Armstrong, J.A. and Horne, R. 1984. Follicular Dendritic Cells and Virus-Like Particles in AIDS-Related Lymphadenopathy. Lancet II:370-372.
119. Tenner-Racz, K., Racz, P., Bofill, M. et al. 1986. HTLV-III/LAV Viral Antigens in Lymph Nodes of Homosexual Men With Persistent Generalized Lymphadenopathy and AIDS. Am. J. Pathol. 123:9-15.
120. Le Tourneau, A., Audouin, J.,Diebold, J. et al. 1986. LAV-like Viral Particles in Lymph Node Germinal Centers in Patients with the Persistent Lymphadenopathy Syndrome and the Acquired Immunodeficiency Syndrome-related Complex. Hum. Pathol. 17:1047-1053.
121. O'Hara, C.J., Groopmen, J.E. and Federman, M. 1988. The Ultrastructural and Immunohistochemical Demonstration of Viral Particles in Lymph Nodes from Human Immunodeficiency Virus-Related Lymphadenopathy Syndromes. Hum. Pathol. 19:545.
122. Culliton, B.J. 1992. The mysterious virus called "isn't". Nature 358:619.
123. Chiodi, F., Albert, J., Olausson, E. et al. 1988. Isolation Frequency of Human Immunodeficiency Virus from Cerebrospinal Fluid and Blood of Patients with Varying Severity of HIV Infection. AIDS Res. Hum. Retroviruses 4:351-358.
124. Salahuddin, S.Z., Groopman, J.E., Markham, P.D. et al. 1984. HTLV-III Symptom-Free Seronegative Persons. Lancet II:1418-1420.
125. Boussin, F.,Rey, F., Dormont, D. et al. 1988. Isolation of HIV in a Seronegative Demented Patient Without Symptoms of Immune Deficiency. Cancer Detect. Prev. 12:257-265.
126. Bayliss, G.J., Jesson, W.J., Evans, B.A. et al. 1989. Isolation of HIV-1 from small volumes of heparinized whole blood. AIDS 3:45-49.
127. Fiore, J.R., Angarano, G., Fico, C. et al. 1990. Cell cultures from small amounts of heparinized whole blood enhance HIV-1 isolation rate. AIDS 4:1295-1296.
128. SchEpbach, J., Jendis, J.B., Bron, C. et al. 1992. False-positive HIV-1 virus cultures using whole blood. AIDS 6:1545-1546.
129. Hart, C., Spira, T., Moore, J. et al. 1988. Direct Detection of HIV RNA Expression in Seropositive Subjects. Lancet II:596-599.
130. Aaronson, S.A., Todaro, G.J. and Scholnick, E.M. 1971. Induction of murine C-type viruses from clonal lines of virus-free BALB/3T3 cells. Science 174:157-159.
131. Hirsch, M.S., Phillips, S.M., Solnik, C. et al. 1972. Activation of Leukemia Viruses by Graft-Versus-Host and Mixed Lymphocyte Reactions In Vitro. Proc. Nat. Acad. Sci. 69:1069-1072.
132. Toyoshima, K. and Vogt, P.K. 1969. Enhancement and Inhibition of Avian Sarcoma Viruses by Polycations and Polyanions. Virol. 38:414-426.
133. Nakamura, N., Sugino, H., Takahara, K. et al. 1991. Endogenous retroviral LTR DNA sequences as markers for individual human chromosomes. Cytogenet. Cell Genet. 57:18-22.
134. Varmus, H. and Brown, P. 1989. Retroviruses, p.53-108. In:Mobile DNA. D.E. Berg, M.M. Howe (Eds.). American Society for Microbiology. Washington, D.C.
135. Weiss, R.A., Friis, R.R., Katz, E. et al. 1971. Induction of Avian Tumor Viruses in Normal Cells by Physical and Chemical Carcinogens. Virol. 46:920-938.
136. Temin, H.M. 1974. On the origin of RNA Tumor Viruses. Harvey Lect. 69:173-197.
137. Wain-Hobson, S. and Myers, G. 1990. Too close for Comfort. Nature 347:18.
138. Genetics of RNA Tumour Viruses. 1973. p.656-699. In:The Molecular Biology of Tumour Viruses. J. Tooze (Ed.). Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
139. Saag, M.S., Hahn, B.H., Gibbons, J. et al. 1988. Extensive Variation of Human Immunodeficiency Virus Type-1 in vivo. Nature 334:440.
140. Marx, J.L. 1988. The AIDS Virus Can Take On Many Guises. Science 241:1039-1040.
141. von Briesen, H., Becker, W.B., Henco, K. et al. 1987. Isolation frequency and growth properties of HIV-Variants:Multiple simultaneous variants in a patient demonstrated by molecular cloning. J. Med. Virol. 23:51-66.
142. Bolton, V., Pedersen, N.C., Higgins, J. et al. 1987. Unique p24 Epitope Marker To Identify Multiple Human Immunodeficiency Virus Variants in Blood from the Same Individuals. J. Clin. Microbiol. 25:1411-1415.
143. Wain-Hobson, S. 1989. HIV genome variability in vivo. AIDS 3:S13-S18.
144. Innocenti, P., Ottmann, M., Morand, P. et al. 1992. HIV-1 Blood Monocytes: Frequency of Detection of Proviral DNA Using PCR in Comparison with the Total CD4 Count. AIDS Res. Hum. Retroviruses 8:261-268.
145. Meyerhans, A., Cheynier, R., Albert, J. et al. 1989. Temporal Fluctuations in HIV quasispecies in vivo are not reflected by sequential HIV isolations.Cell 58:901-910.
146. Robey, W.G., Nara, P.L., Poore, C.M. et al. 1987. Rapid Assessment of Relationships Among HIV Isolates by Oligopeptide Analyses of External Envelope Glycoproteins. AIDS Res. Hum. Retroviruses 3:401-408.
147. Van Voorhis, B.J., Martinez, A., Mayer, K. and Anderson, D.J. 1991. Detection of human immunodeficiency virus type 1 in semen from seropositive men using culture and polymerase chain reaction deoxyribonucleic acid amplification techniques. Fertil. Steril. 55:588-594.
148. Shaw, G.M., Hahn, B.H., Suresh, K.A. et al. 1984. Molecular Characterization of Human T-Cell Leukemia (Lymphotropic) Virus Type III in the Acquired Immune Deficiency Syndrome. Science 226:1165-1171.
149. Simmonds, P., Balfe, P., Peutherer, J.F. et al.1990. Human Immunodeficiency Virus-Infected Individuals Contain Provirus in Small Numbers of Peripheral Mononuclear Cells and at Low Copy Numbers. J. Virol. 64:864-872.
150. Defer, C., Agut, H., and Garbarg-Chenon, A. 1992. Multicentre quality control of polymerase chain reaction for detection of HIV DNA. AIDS 6:659-663.
151. Becker, J.L., Hazan, U., Nugeyre, M. T. et al. 1986. Infection of insect lines by HIV, agent of AIDS, and evidence for HIV proviral DNA in insects from Central Africa. C. R. Acad. Sci. Paris. 300:303-306.
152. Arya, S.K., Gallo, R.C., Hahn, B.H. et al. 1984. Homology of Genome of AIDS-Associated Virus with Genomes of Human T-cell Leukemia Viruses. Science 225:927-930.
153. Mager, D.L. and Freeman, J.D..1987. Human Endogenous Retroviruslike Genome with Type C pol Sequences and gag Sequences Related to Human T-Cell Lymphotropic Viruses. J. Virol. 61:4060-4066.
154 Banki, K., Maceda, J., Hurley, E. et al. 1992. Human T-cell lymphotropic virus (HTLV)-related endogenous sequence, HRES-1 encodes a 28-kDa protein:A possible autoantigen for HTLV-I gag-reactive autoantibodies. Proc. Natl. Acad. Sci. 89:1939-1943.
155. Webster, A.D.B., Dalgleish, A.G., Beattie, R. et al. 1986. Isolation of retroviruses from two patients with "Common Variable" Hypogammaglobulinaemia. Lancet I:581-582.
156. Ciampolillo, A., Marini, V. and Buscema, M. 1989. Retrovirus-like Sequences in Graves' Disease:Implications for Human Autoimmunity. Lancet I:1096-1100.
157. Horwitz, M.S., Boyce-Jacino, M.T. and Faras, A.J. 1992. Novel Human Endogenous Sequences Related to Human Immunodeficiency Virus Type 1. J. Virol. 66:2170-2179.
158. Farzadegan, H., Polis, M., Wolinsky, S.M. et al. 1988. Loss of Human Immunodeficiency Virus Type 1 (HIV-1) Antibodies with Evidence of Viral Infection in Asymptomatic Homosexual Men. Ann. Int. Med. 108:785-790.
159. Horsburgh, C.R., Ou, C.Y., Holmberg, S.D. et al. 1989. Human Immunodeficiency Virus Type 1 Infection in Homosexual Men who remain Seronegative for prolonged periods. NEJM 321:1678-1680.
160. Mikovits, J.A., Lohrey, N., Schuloff, R., Ruscetti, F. 1991. Immune Activation of latent HIV-1 expression in monocyte/macrophages, p151. In:Vol. I, Abstracts VII International Conference on AIDS,Florence.
161. Vartanian, J.P., Meyerhans, A., Henry, M. and Wain-Hobson, S. 1992. High-resolution structure of an HIV-1 quasispecies:identification of novel coding sequences. AIDS 6:1095-1098.